
Аннотация
По сравнению с королевой или рабочими особями, биология трутней медоносной пчелы Apis mellifera L. мало изучена. Имеющаяся информация об активности трутней основана в основном на прямых наблюдениях в течение ограниченного периода времени и в ограниченное время суток. Полные регистры летной активности трутней медоносных пчел отсутствуют. Мы изучили активность трутней A. mellifera в течение всей их жизни весной и летом, используя оптический счетчик пчел у входа в улей. Трутни были активны во второй половине дня, причем большинство полетов происходило между 14:00 и 18:00. Короткие ориентационные полеты совершались в возрасте 6-9 дней, а более длительные брачные полеты продолжительностью 30 мин совершались с 21-дневного возраста весной и с 13-дневного возраста летом. Наши регистры показывают, что 50 и 80% трутней оставались верными своей колонии (не дрейфовали) весной и летом, соответственно. Настоящее исследование подтверждает существующую информацию, но также раскрывает неизвестные аспекты биологии трутней.
1 Введение
С девятнадцатого века медоносная пчела является очень привлекательной моделью для социобиологии и поведенческих наук. Apis mellifera L. была широко изучена, и среди различных каст колонии ученые сосредоточились в основном на королеве и рабочих и меньше на поведении и биологии трутней. Несмотря на то, что трутни не участвуют в сборе пищи или уходе за выводком, существуют сложные взаимодействия, в которых рабочие стимулируют трутней к трофаллаксису вибрационными сигналами (Slone et al. 2012; Goins and Schneider 2013). Кроме того, известно, что тепло, выделяемое трутнями, участвует в терморегуляции гнезда (Harrison 1987; Kovac et al. 2009). Дроны привлекают внимание в основном из-за их удивительного брачного поведения вне гнезда. Зрелые трутни покидают гнездо и летят в определенные места скопления трутней (DCAs), чтобы спариться с королевой. На высоте 15-40 м в воздухе (Ruttner 1966), в DCA, десятки тысяч трутней собираются и ждут королеву. Копуляция происходит на крыле, и трутни погибают вскоре после спаривания (Koeniger et al. 2005a). Только несколько исследований описывают летную активность и продолжительность жизни трутней медоносной пчелы (Howell and Usinger 1933; Oertel 1956; Ruttner 1966; Witherell 1971; Hellmich 1991; Koeniger et al. 2005a; Koeniger et al. 2005b). Молодые особи совершают ориентационные полеты вокруг гнезда, прежде чем достичь половой зрелости несколько дней спустя (Howell and Usinger 1933; Koeniger et al. 2005a). Ориентационные полеты — это специфическое поведение пчел, которое заключается в зависании взад и вперед у входа в улей, а затем в подъеме по спирали. Во время таких ориентационных полетов пчелы изучают особенности ландшафта, которые позволяют им вернуться в улей (хоминг) (Capaldi and Dyer 1999; Capaldi et al. 2000), а опыт полетов улучшает показатели возвращения (Hayashi et al. 2017). По данным Hellmich et al. (1991), время вылета зрелых трутней составляет около 16:00, при этом различия между незрелыми и зрелыми трутнями незначительны. При благоприятных погодных условиях трутни могут совершать несколько брачных полетов в течение одного дня, оставаясь в DCA до 30 минут, прежде чем вернуться в улей для кормления (Gary 1992). Однако имеющаяся информация о полетном поведении трутней обычно основана на прямых наблюдениях, в течение ограниченного периода времени и только в ограниченные временные интервалы (Burgett 1974; Fukuda and Ohtani 1977; Hellmich et al. 1991; Duay et al. 2002; Rueppell et al. 2005; Neves et al. 2011). Кроме того, эти наблюдения не позволяют идентифицировать трутней по отдельности (Burgett 1974; Rueppell et al. 2005), и трудно определить точное количество трутней, вылетающих из улья или прибывающих в него, особенно когда сотни трутней летают одновременно (Rinderer et al. 1993). Кроме того, время полета может меняться в зависимости от погодных условий (Hellmich et al. 1991). Таким образом, информация, которую можно получить при непосредственном наблюдении за поведением трутней, ограничена.
Дроны могут перелетать из одной колонии в другую, и обычно они легко принимаются в чужие колонии в весенний период. Дрейф трутней считается обычным явлением на пасеках (Currie and Jay 1991; Moritz and Neumann 1996; Neumann et al. 2000), но эмпирические доказательства их верности собственной колонии отсутствуют. Чтобы лучше понять поведение трутней медоносной пчелы и поведенческое развитие летной активности, мы использовали оптический счетчик пчел, способный регистрировать выходы и входы трутней, помеченных индивидуальными метками. Мы сообщаем о частоте и продолжительности полетов, а также о дрейфе.
2 Материал и методы
Пустые рамки для трутней были введены в колонию с сильной королевой, чтобы получить новых трутней того же возраста. Отложенные королевой яйца развивались в трутней через 25 дней. Сто появившихся трутней (в возрасте 0-12 ч) из той же колонии были индивидуально помечены отличительной меткой с номерами от 0 до 99, приклеенной к грудной клетке в вертикальном положении (по направлению к голове) (рис. 1). Метка позволяла регистрировать активность каждого трутня. До этого исследования мы подтвердили, что метки не влияют на выживаемость трутней в лабораторных условиях (данные не показаны). Трутни с метками были введены в колонию A. mellifera, содержащуюся в улье Дадана в INRA-Avignon (Франция). Улей состоял из 5 рамок и около 10 000 рабочих особей. Пасека в INRA-Avignon состоит примерно из 30 полноразмерных колоний и около 20 пятирамочных колоний. Экспериментальный улей был размещен на расстоянии 20 м от других колоний и между густыми кустами, чтобы избежать дрейфа.

Активность меченых трутней, выходящих из улья и входящих в него, регистрировалась с помощью оптического счетчика пчел, основанного на системе распознавания видеокамер и устройстве прохода, через который трутень должен пройти, чтобы войти или выйти из улья (Le Conte and Crauser 2006; Dussaubat et al. 2013; Alaux et al. 2014; Prado et al. 2019). Система постоянно регистрировала активность выхода и входа помеченных трутней.
Оптический счетчик пчел состоял из камеры высокого разрешения, которая следила за входом в улей, и собственного программного обеспечения для анализа изображений, которое обнаруживало и регистрировало метку. Программное обеспечение способно определять направление движения метки и отличать выходы от входов. Идентификация номеров осуществляется с помощью оптического распознавания символов (OCR), при этом голова считается поднятой. Для входящего беспилотника изображение поворачивается на 180° перед регистрацией. Устройство прохода состоит из пяти проходов, закрытых плексигласом, а система освещения позволяет камере вести запись. Изображения анализируются из двух зон обнаружения, и пчелы учитываются только в том случае, если они пересекают обе зоны. Это ограничивает анализ пчелами, которые действительно покидают или возвращаются в улей.
Полеты дронов определялись как последовательность выхода-входа для определенного идентификатора дрона. Непарные выходы или входы, вызванные незамеченными проходами, не учитывались (6% данных). Одновременно оптический счетчик регистрирует глобальную активность входа-выхода не меченых рабочих в улье. Для данного исследования была выбрана колония из 5 рамок, чтобы уменьшить интенсивность движения на проходе и повысить точность записи.
Два повторных исследования были проведены в течение 90 дней на одном и том же улье, оба со 100 трутнями. Первая репликация была проведена в мае-июне (весной), а вторая — в июне-июле (летом). В обоих случаях данные собирались до тех пор, пока последний трутень не покинул улей.
Погодные данные (температура, скорость ветра, дождь) и время восхода и захода солнца были получены в течение двух периодов исследования на сайтах http://fr.weather.com/climate/sunRiseSunSet-Avignon-FRXX0270 и http://www.infoclimat.fr/tableaux-station-meteo-avignon-07563. Поскольку пик активности трутней приходился на 16:00 (см. раздел 3), коэффициенты корреляции (r Пирсона) между активностью трутней (минуты, проведенные вне улья) и метеорологическими условиями в 16:00-17:00 оценивались с помощью корреляционных тестов с использованием функции cor.test в R.
Для сравнения активности трутней (количество полетов и общее количество минут, проведенных вне улья в день) между весенним и летним периодами использовали U-тест Манна-Уитни, реализованный в R.
3 Результаты
3.1 Возраст первого и последнего полетов
Полная запись полетов была получена для 90 и 86 трутней в весеннем и летнем опытах, соответственно. Весной 74% особей совершили первый полет в возрасте 6 дней, а летом 51,7% совершили первый полет в возрасте 7 дней. Средний возраст при первом полете составил 5,6 дня (± 0,9 SD) и 7,2 дня (± 0,5 SD) для весеннего и летнего экспериментов, соответственно.
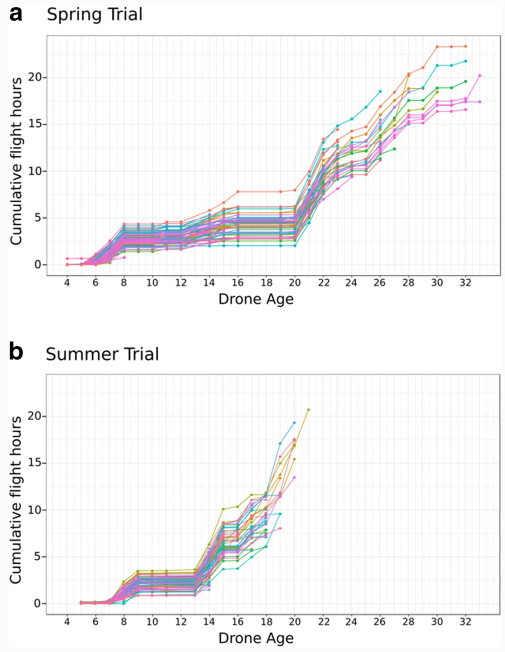
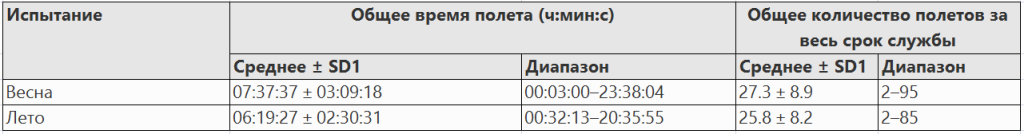
Средний возраст, в котором дроны совершили свой последний полет весной, составил 17,9 дней (± 4,4 SD). Летом последний полет произошел значительно раньше, чем весной (U = 3347, N1 = 90, N2 = 86, P = 0,013), в среднем через 15,2 дня (± 1,8 SD) (Таблица I). Максимальная продолжительность жизни, наблюдаемая весной и летом, составила 33 и 21 день соответственно (табл. I).

3.2 Ориентационные и брачные полеты
Как сообщают другие авторы (Witherell 1971; Currie 1987), в наших экспериментах дроны совершали два типа полетов. Короткие полеты (ориентация), выполняемые молодыми дронами, и более длительные полеты (интерпретируемые как брачные полеты, когда дроны посещают DCA), выполняемые в более позднем возрасте. В обоих экспериментах дроны совершали короткие полеты продолжительностью около 15 минут в возрасте 6-8 дней в весеннем эксперименте (15,2 мин ± 10,1 SD) и в возрасте 7-9 дней в летнем (14,8 мин ± 11,8 SD). Весной, начиная с возраста 21 день, выполнялись более длительные брачные полеты продолжительностью 31,5 мин (± 11,4 SD). Во время летних испытаний трутни совершали длительные брачные полеты продолжительностью 27,4 мин (± 14,5 SD), начиная с 13-дневного возраста. Самый продолжительный индивидуальный полет трутней произошел весной в возрасте 28 дней (2 ч 51 мин) и летом в возрасте 16 дней (1 ч 45 мин). В обоих случаях это произошло в последний день полета.
Между ориентационным и брачным полетами, несмотря на благоприятные погодные условия, мы наблюдали период очень низкой активности в течение 12 и 5 дней весной и летом, соответственно. В этот период трутни редко покидали улей (рис. 2).
Время, проведенное в ДКА (оцененное как общее время брачных полетов), составило в среднем 510,9 мин (± 268,5 SD) и 327,3 мин (± 252,6 SD) весной и летом, соответственно. Мы обнаружили положительную связь между общим временем, затраченным на выполнение ориентационных полетов, и общим временем, проведенным в DCA (r = 0,31, P = 0,003, и r = 0,35, P = 0,001, весной и летом, соответственно, рис. 3).

3.3 Количество полетов в день, летные дни, полеты в течение жизни и общее время полета
Дроны выполняли 3,3 (± 1,4) и 2,7 (± 1,8) ориентационных полетов в день весной и летом, соответственно. Это составило в общей сложности 8,8 (± 3,3) и 8,1 (± 3,1) ориентационных полетов за жизнь трутня, после чего они в основном оставались внутри улья до 21 дня весной и 14 дней летом, когда они начинали совершать более длительные полеты (предположительно брачные полеты, во время которых трутни посещают ДКА). Весной трутни совершали в среднем 4,3 ± 2,6 брачных полета в день. Эти брачные полеты совершались в среднем в течение 5,1 ± 3,5 дней, не следующих друг за другом, что в общей сложности составляет 22 ± 14 брачных полетов в жизни дрона весной. В летнем испытании дроны летали больше раз в день (U = 18 792, N1 = 224, N2 = 212, P = 0,0001), но в меньшее число дней (U = 2016, N1 = 46, N2 = 70, P = 0,02078; Таблица II). Летом они совершали 5,7 брачных полетов в день только в течение 3,4 ± 2 дней, не следующих друг за другом, в общей сложности 20 ± 19 брачных полетов (рис. 4a).

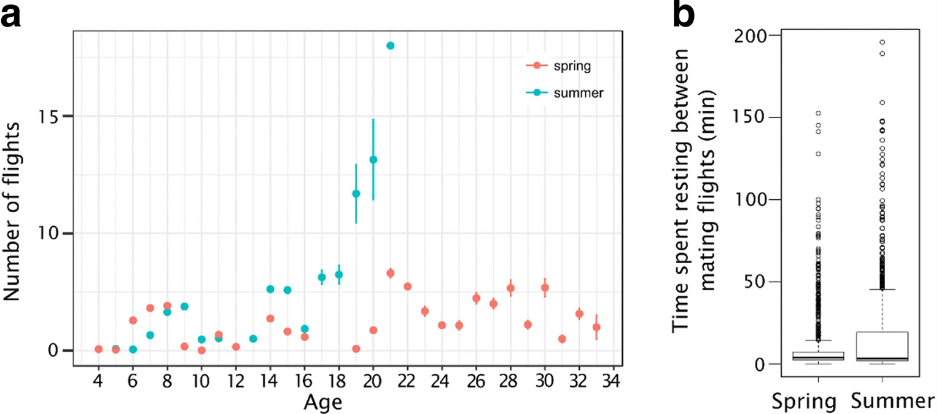
Между брачными полетами трутни восстанавливают силы, питаясь медом (Witherel 1971; Currie 1987). В нашем исследовании трутни провели 10,7 ± 19,1 и 14,7 ± 24,7 мин (среднее ± SD) между брачными полетами внутри улья весной и летом, соответственно (медиана 3,8 и 3,3 мин) (рис. 4b).
Количество общих полетов на одного трутня (ориентация + спаривание) варьировало от 1 до 106, в среднем 27,3 ± 20,06 и 27,4 ± 21,5 полетов весной и летом, соответственно. Высокая изменчивость общего числа полетов частично объясняется процентом дронов, которые не прожили достаточно долго, чтобы совершить брачные полеты; 48% и 19% весной и летом, соответственно, не совершили ни одного брачного полета. Общее количество полетов на одного трутня не отличалось между двумя оцениваемыми сезонами (U = 3890, N1 = 90, N2 = 86, P = 0,954). Не отличалось и суммарное время полета (U = 4241, N1 = 90, N2 = 86, P = 0,2728): в среднем 7 ч 37 мин 37 с весной и 6 ч 19 мин 27 с летом (табл. III).
3.4 Период полета
В обоих испытаниях самые ранние зарегистрированные полеты половозрелых дронов были в 7:00, хотя это случалось очень редко (Рисунок 5a, c). В целом, дроны начинали свои полеты во второй половине дня (Рисунок 5a, c), и большинство ежедневных полетов происходило между 14:00 и 18:00, с заметным пиком активности в 16:00 (Рисунок 5b, d). Летом продолжительность времени активности на открытом воздухе была на 3 ч больше, чем весной (U = 4724, N1 = 237, N2 = 240, P = 0,0135), что означает, что некоторые трутни начинали летать раньше и позже возвращались в свой улей.
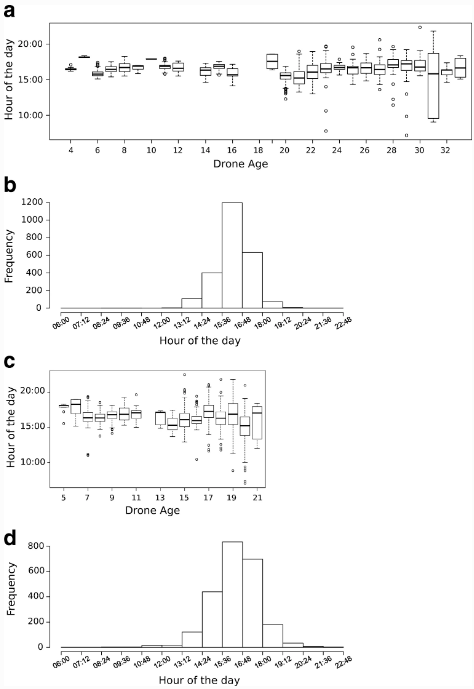
Низкие, но значимые корреляции наблюдались между восходом солнца и ежедневным временем начала полетов дронов (r = 0,394, P = 0,0335, и r = — 0,611, P = 0,0104, для испытаний 1 и 2, соответственно), а также между продолжительностью дня и временем начала полетов дронов (r = — 0,401, P = 0,0303, и r = 0,687, P = 0,0024, для испытаний 1 и 2, соответственно).
Дроны не были постоянно активны на протяжении всей своей жизни. Напротив, мы наблюдали пики активности (рис. 6). Первый пик активности наблюдался почти в одном и том же возрасте (6-9 дней) в обоих опытах, что соответствует ориентационным полетам (Burgett 1974; Capaldi and Dyer 1999; Capaldi et al. 2000). Более поздние всплески активности происходили по достижении трутнями возраста половой зрелости и были связаны преимущественно с благоприятными климатическими условиями, позволяющими совершать брачные полеты (рис. 6). Однако, несмотря на благоприятные погодные условия, в некоторых случаях трутни оставались в улье после двух дней брачных полетов.
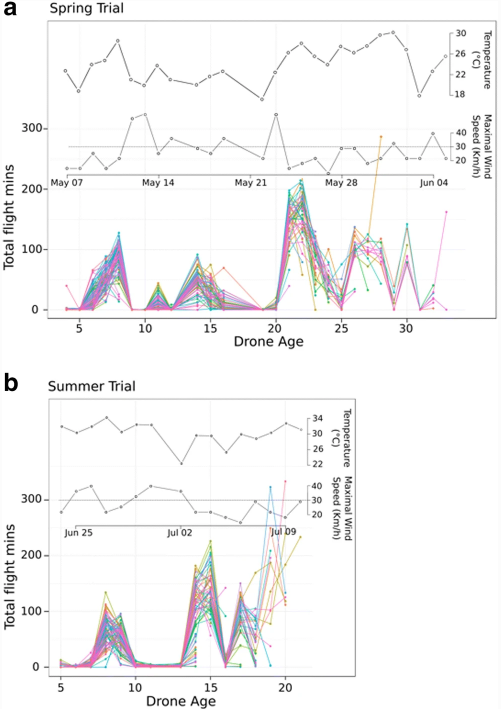
3.5 Влияние погодных условий на полеты трутней
По нашим наблюдениям, трутни редко вылетали из улья, если скорость ветра достигала 30 км/ч или шел дождь (только 2 дождя было весной и ни одного летом). Однако несколько трутней смогли летать в дни, когда ветер достигал 35 км/ч. Эти дроны совершили несколько коротких полетов (от 30 с до 7 мин) в течение дня, в то время как большинство дронов ждали, пока скорость ветра опустится ниже 30 км/ч, чтобы начать свои полеты (рис. 6). Температура в 16:00 коррелировала с активностью дронов весной, но не летом, когда дневная температура оставалась выше 19 °C (Таблица IV). Летом трутни продолжали совершать брачные полеты на час позже после полудня, чем весной (рис. 5).

3.6 Возвращение в улей и дрейф
Данные оптического счетчика использовались для оценки количества трутней, улетевших в другие колонии, путем оценки количества трутней, которые провели одну или несколько ночей в другом месте, прежде чем вернуться. Этот показатель недооценивает дрейф трутней, поскольку он не учитывает трутней, которые постоянно дрейфуют в другие колонии. Во время весеннего испытания 7 трутней (7,7%) ушли в другие колонии и вернулись через 1-13 дней, а 46 (50,5%) остались верны своей колонии. Три трутня погибли внутри улья (последний раз обнаружены входящими в улей), а 35 (38,5%) могли уйти в другие колонии и не вернуться, или погибнуть вне улья до начала брачных полетов. Во время летнего испытания три трутня (3,5%) ушли в другие колонии и вернулись через 1-6 дней, а 70 (80%) остались верны своей колонии до начала брачных полетов (14 дней). Четыре трутня погибли внутри улья, а 10 (11,5%) могли уйти в другие колонии и не вернуться, или погибнуть вне улья, не достигнув половой зрелости (рис. 7).

3.7 Полеты дронов и рабочих пчел
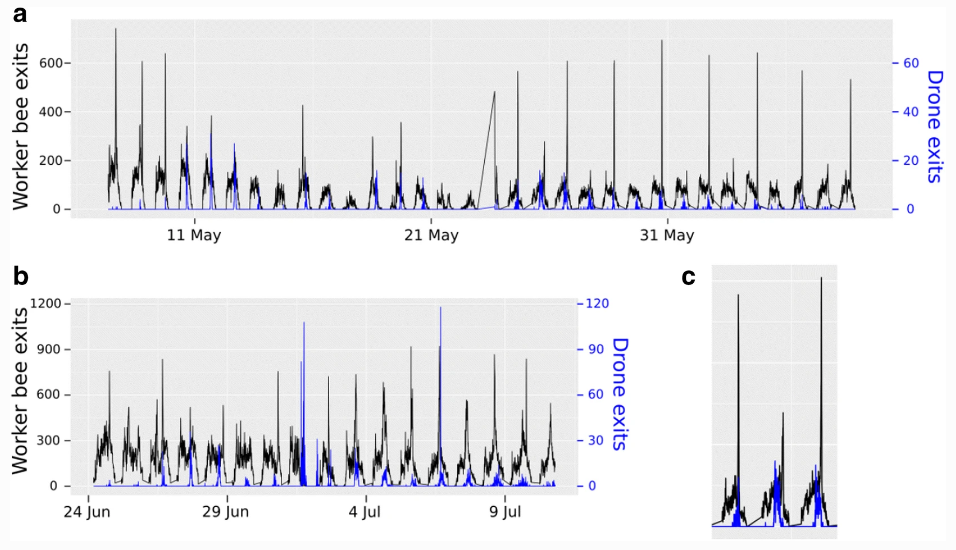
Мы наблюдали ежедневный пик активности в полетах рабочих и трутней, но не обнаружили четкой взаимосвязи между ежедневным пиком активности трутней (16:00) и послеобеденным пиком рабочих пчел. В то время как самцы пчел оставались постоянными в своем пике активности в 16:00 (Рисунок 5), пик активности рабочих пчел был разным каждый день и, похоже, был более чувствителен к изменениям погодных условий (Рисунок 8).
4 Обсуждение
Мы зарегистрировали прижизненную летную активность трутней медоносной пчелы, подтвердив имеющиеся данные (Burgett 1974; Hellmich et al. 1991; Neves et al. 2011), но также предоставив новую информацию в отношении возраста трутней и метеорологических условий. Дроны начали выполнять ориентировочные полеты в возрасте 6-7 дней, как сообщалось ранее (Tozetto et al. 1997; Colonello-Frattini and Hartfelder 2009). Возраст последнего трутня, покинувшего улей (21 и 33 дня), также соответствовал ранее опубликованным данным (Winston 1987).
В данном исследовании продолжительность жизни оценивалась как время последней записи. Мы наблюдали более долгоживущих трутней весной по сравнению с летом. Фукуда и Охтани (1977) также обнаружили разницу между продолжительностью жизни трутней в разные сезоны (осень и лето), причем осенние трутни жили дольше. В обоих случаях жизнь пчелиных самцов была короче летом, что может быть связано с более высокими температурами и количеством брачных полетов в день. В нашем исследовании средняя температура весной была на 5,2 °C ниже, чем летом. Летом период дня, в течение которого трутни были активны, увеличился на 3 часа, а количество брачных полетов возросло с 4,3 до 5,7 часов в день. Роуэлл и др. (1986) указывают, что высокая смертность летом, вероятно, связана как с высоким уровнем летной активности, так и с метеорологическими экстремальными условиями, связанными с ресурсным стрессом для колонии. В нашем летнем исследовании продолжительность жизни достигла всего 15,2 дня. Это значение сходно с результатами Фукуды и Охтани (1977), которые определяли выживаемость путем подсчета количества помеченных трутней, вернувшихся в улей. Другие исследования, однако, показывают, что продолжительность жизни летних трутней составляет от 21,2 до 23,5 дней (Witherell 1972).
В обоих опытах мы наблюдали, что начало брачных полетов было связано с повышением дневной температуры (рис. 6). Интересно, что в обоих опытах после двух дней совершения брачных полетов трутни оставались в улье, чтобы продолжить полет через один или два дня, которые показаны на рисунке 6 как пики активности. Эта пауза в активности, похоже, не связана с климатическими условиями в летнем испытании, поскольку погодные условия были хорошими для полетов. Следовательно, пауза в активности может отражать физиологическое ограничение.
Мы обнаружили слабую, но значительную положительную связь между временем, затраченным на выполнение ориентационных полетов, и временем, затраченным на брачные полеты. Дроны, которые тратили больше времени на ориентацию, также проводили больше времени в DCA (по оценке времени брачных полетов). Эта взаимосвязь может отражать большую летную способность некоторых особей или важность обучения в ходе ориентационных полетов (Capaldi and Dyer 1999; Capaldi et al. 2000). Манипулятивные исследования, ограничивающие время ориентации дронов, могут пролить свет на роль обучения в работе дронов.
Ветер оказывает важное влияние на полет трутней. В то время как мы обнаружили, что большинство трутней не летали в дни, когда ветер был > 30 км/ч, независимо от их возраста, Ленски и Демптер (1985) наблюдали задержку полетов маток и трутней при ветре > 14 км/ч. Аналогично, Дуай и др. (2002) указали, что самцы пчел не летают, когда ветер превышает 15 км/ч. В данном исследовании некоторые трутни совершили несколько коротких полетов при средней скорости ветра до 35 км/ч, что говорит о том, что матка может быть более чувствительной к ветру, чем трутни, и что трутни с лучшими показателями могут быть более конкурентоспособными при спаривании с маткой. Мы также наблюдали, что после ветреного дня, когда трутни не покидали улей, они совершали короткий полет рано утром на следующий день. Короткие полеты в ветреные дни или рано утром могут быть гигиеническими полетами (Page and Peng 2001).
Мы обнаружили, что активность трутней не происходит исключительно во второй половине дня, как предполагалось в предыдущих исследованиях (Burguett 1974; Fukuda and Ohtani 1977; Hellmich et al. 1991; Duay et al. 2002; Rueppell et al. 2005). В зависимости от погодных условий некоторые самцы летали рано утром (7-9 ч) или поздно вечером (19-22 ч). Мы также наблюдали, что летние трутни начинали и заканчивали свои ежедневные полеты позже, чем весенние. Королевы A. mellifera совершают брачные полеты между 12:00 и 17:00 с максимальной летной активностью между 13:00 и 16:00 (Heidinger et al. 2014). Ленски и Демтер (1985) заметили, что продолжительность полетов королевы сокращается при температурах 15 и 20 °C. Это было подтверждено Heidinger et al. Это было подтверждено Хайдингером и др. (2014), которые сообщили, что при высоких температурах королевы летают реже, но в течение более длительных периодов времени по сравнению с низкими температурами. В идеале, организмы должны распределять свою активность по времени таким образом, чтобы максимизировать физическую нагрузку; следовательно, пик активности трутней в 16:00 может отражать эту оптимизацию.
Дроны могут быть приняты в разные ульи, когда условия благоприятны (погода и наличие пищи) (Galindo-Cardona et al. 2015). По данным Jaffé и Moritz (2010), дрейф трутней A. mellifera может достигать 10% трутней в улье. Это значение согласуется с нашими наблюдениями, поскольку 7 и 4% трутней были подтвержденными дрифтерами (трутни, которые вернулись после того, как провели одну или несколько ночей вне улья). Кроме того, часть трутней, которые покинули улей и не вернулись, могли потенциально переместиться в другие колонии (эти трутни считались неучтенными на рисунке 7) (Fukuda and Ohtani 1977). В нашем исследовании 50 и 77% весенних и летних трутней, соответственно, оставались верными своей колонии до начала брачных полетов.
Как и в работе Woyke et al. (2001) для Apis dorsata, мы также обнаружили отрицательную корреляцию между временем восхода солнца и началом полета дронов. Однако в нашем исследовании уровень корреляции был ниже, возможно, из-за меньшей амплитуды в изменении времени восхода солнца. В то время как авторы наблюдали разницу в один час между первой и последней датой в своем исследовании, в настоящем исследовании эта разница составила всего 39 минут в летнем испытании.
Сообщается, что беспилотники летают со скоростью 9-11,5 км/ч (Gmeinbauer and Crailsheim, 1993; Koeniger et al. 2005a). Сообщалось, что ДКА пролетают в непосредственной близости от пасек, максимальное расстояние до которых составляет 7 км (Ruttner and Ruttner, 1972). Учитывая, что мы зарегистрировали среднюю продолжительность полета 30 мин, большинство трутней, должно быть, посещали места скопления на расстоянии 2-4 км от улья. Очень немногие полеты превышали 60 мин, а самый длинный полет, который мы зарегистрировали, длился 171 мин. Эти редкие длительные полеты могли быть связаны с тем, что трутни посещали ДКП на более дальние расстояния, оставались в ДКП в течение длительного времени или перелетали в другую колонию между двумя брачными полетами (Galindo-Cardona et al. 2015).
Оптические счетчики пчел — полезные инструменты, позволяющие изучать поведение сотен отдельных пчел. Мы предоставили подробную информацию о полетном поведении и наружной активности трутней, касающуюся времени суток их активности, а также количества и продолжительности полетов. Мы заметили, что большинство трутней сохраняют верность своей колонии до начала брачных полетов.
Дроны — социальные существа, и их брачную деятельность следует рассматривать в связи с колонией (Ruttner 1966). В данном исследовании мы использовали небольшой 5-рамочный улей; небольшой размер колонии мог повлиять на активность трутней. Для подтверждения наших выводов необходимы будущие исследования с использованием полноразмерных колоний.
Ссылки
1.Alaux, C., Crauser, D., Pioz, M., Saulnier, C., Le Conte, Y. (2014) Parasitic and immune-modulation of flight activity in honey bees tracked with optical counters. J. Exp. Biol. 217, 3416–3424. Article Google Scholar
2.Burgett, M. (1974) Drone honey bee flight from clustered swarms. Ann. Entomol. Soc. Am.67, 683–684.
3.Capaldi, E.A., Dyer, F.C. (1999) The role of orientation flights on homing performance in honeybees. J. Exp. Biol. 202, 1655–1666. CAS PubMed Google Scholar
4.Capaldi, E.A., Smith, A.D., Osborne, J.L., Fahbach, S.E., Farris, S.M., Reynolds, D.R., Edwards, A.S., Martin, A., Robinson, G.E., Poppy, G.M., Riley, J.R. (2000) Ontogeny of orientation flight in the honeybee revealed by harmonic radar. Nature 403, 537–540.
Article CAS Google Scholar
5.Colonello-Frattini, N., Hartfelder, K. (2009) Differential gene expression profiling in mucus glands of honey bee (Apis mellifera) drones during sexual maturation. Apidologie 10, 481–495.
Article Google Scholar
6.Currie, R W. (1987) The biology and behaviour of drones. Bee World 68, 129–143.
Article Google Scholar
7.Currie, R.W. and Jay, S.C. (1991) Drifting behaviour of drone honey bees (Apis mellifera L) in commercial apiaries. J. Apic. Res. 30, 61–68.
Article Google Scholar
8.Duay, P., De Jong, D., Engels, W. (2002) Decreased flight performance and sperm production in drones of the honey bee (Apis mellifera) slightly infested by Varroa destructor mites during pupal development. Genet. Mol. Res. 1, 227–232.
PubMed Google Scholar
9.Dussaubat, C., Maisonnasse, A., Crauser, D., Beslay, D., Costagliola, G., Soubeyrand, S., Kretzchmar, A., Le Conte, Y. (2013) Flight behavior and pheromone changes associated to Nosema ceranae infection of honey bee workers (Apis mellifera) in field conditions. J. Invert.Pathol. 113, 42–51.
Article CAS Google Scholar
10.Fukuda, H., Ohtani, T. (1977). Survival and life span of drone honeybees. Res. Popul. Ecol.19, 51–68.
11.Gary, N.E. (1992) Activities and behaviour of honey bees. In: Graham JM (Ed.), The hive and the honey bee. Dadant & Sons Inc., Hamilton, pp. 269–372. Google Scholar
12.Goins, A., Schneider, S.S. (2013) Drone “quality” and caste interactions in the honey bee, Apis mellifera L. Insectes Soc. 60, 453–461.
13.Gmeinbauer, R., Crailsheim, K. (1993) Glucose utilization during flight of honeybee (Apis mellifera) workers, drones and queens. J. Insect Physiol.39, 959–967.
14.Harrison, J.M. (1987) Roles of individual honeybee workers and drones on colonial thermogenesis. J. Exp. Biol.129, 53–61.
15.Hayashi, S., Farkhary S., Takata, M., Satoh, T., Koyama, S. (2017) Return of drones: Flight experience improves returning performance in honeybee drones. J. Insect Behav. 30(3), 237–246.
Article Google Scholar
16.Heidinger, I.M.M., Meixner, M.D., Berg, S., Büchler, R. (2014) Observation of the mating behavior of honey bee (Apis mellifera L.) queens using radio-frequency identification (RFID): factors influencing the duration and frequency of nuptial flights. Insects 5, 513–527.
Article Google Scholar
17.Hellmich, R.L., Rinderer, T.E., Danka, R.G., Collins, A.M., Boykin, D.L. (1991) Flight times of Africanized and European honey bee drones (Hymenoptera: Apidae). J. Econ. Entomol.84, 61–64.
18.Howell, D.E., Usinger, R.L. (1933) Observations on the flight and length of life of drone bees. Ann. Entomol. Soc. Am.26, 239–246.
19.Jaffé, R.,Moritz, R. (2010) Mating flights select for symmetry in honeybee drones (Apis mellifera). Naturwissenschaften 97, 337–343. Article Google Scholar
20.Koeniger, N., Koeniger, G., Gries, M., Tingek, S. (2005a) Drone competition at drone congregation areas in four Apis species. Apidologie 36, 211–221. Article Google Scholar
21.Koeniger, N., Koeniger, G., Pechhacker, H. (2005b) The nearer the better? Drones (Apis mellifera) prefer nearer drone congregation areas. Insect Soc.52, 31–35.
22.Kovac, H., Stabentheiner, A., Brodschneider, R. (2009) Contribution of honeybee drones of different age to colonial thermoregulation. Apidologie,40, 82–95. Article Google Scholar
23.Le Conte, Y., Crauser, D. (2006) Vers de nouveaux systèmes de comptages automatiques d’abeilles. Bull. Tech. Apic.33, 23–30.
24.Lensky, Y., Demter, U. (1985) Mating flight of the queen honeybee (Apis mellifera) in a subtropical climate. Comp. Biochem. Physiol.81A, 229–241.
25.Moritz, R.F.A., Neumann, P. (1996) Genetic analysis of the drifting of drones in Apis mellifera using multilocus DNA fingerprinting. Ethology 102(7), 580–590. Google Scholar
26.Neumann, P., Moritz, R.F.A., Mautz, D. (2000) Colony evaluation is not affected by drifting of drone and worker honeybees (Apis mellifera L.) at a performance testing apiary. Apidologie 31, 67–79. Article Google Scholar
27.Neves, E., Faita, M., de Oliveira Gaia, L, Vieira Alves Júnior, V., Antonialli-Junior, F. (2011) Influence of Climate Factors on Flight Activity of Drones of Apis mellifera (Hymenoptera: Apidae). Sociobiology57,107–111. Google Scholar
28.Oertel, E (1956) Observation on the flight of drone honey bees. Ann. Entomol. Soc.Am. 49, 497–500. Article Google Scholar
29.Page, R., Peng, C. (2001) Aging and development in social insects with emphasis on the honey bee, Apis mellifera L. Exp. Gerontol. 36, 695–711.
30.Prado, A., Pioz, M., Vidau, C., Requier, F., Jury, M., Crauser, D., Brunet, J-L., Le Conte, Y., Alaux, C. (2019) Exposure to pollen-bound pesticide mixtures induces longer-lived but less efficient honey bees. Sci. Total Environ. 650, 1250–1260. Article CAS Google Scholar
31.Rinderer, T., Oldroyd B, Wonsiri, S., Allen Sylvester, H., De Guzman L, Potichot, S. Sheppard, W., Buckmann, S (1993) Time of drone flight in four honey bee species in south-eastern Thailand. J. Apic. Res. 32 (1), 27–33. Article Google Scholar
32.Rueppell, O., Fondrk, M.K. and Page, R.E. (2005) Biodemographic analysis of male honey bee mortality. Aging cell 4, 13–19. Article CAS Google Scholar
33.Ruttner, H. (1966) The Life and Flight Activity of Drones. Bee World 47(3), 93–100. Article Google Scholar
34.Ruttner, H., Ruttner, F. (1972). Untersuchungen über die Flugaktivität und das Paarungsverhalten der Drohnen. V.-Drohnensammelplätze und Paarungsdistanz. Apidologie, 3, 203–232. Article Google Scholar
35.Rowell, G.A., Taylor, O.R., Locke, S.J. (1986) Variation among commercial honey bee stocks. Apidologie 17(2), 137–158. Article Google Scholar
36.Slone, J.D., Stout, T.L., Huang, Z.Y., Schneider, S.S. (2012) The influence of drone physical condition on the likelihood of receiving vibration signals from worker honey bees, Apis mellifera. Insectes Soc.59, 101–107.
37.Tozetto, S., Rachinsky, A., Engels, W. (1997) Juvenile hormone promotes flight activity in drones (Apis mellifera carnica). Apidologie 28, 77–84. Article Google Scholar
38.Winston, M. (1987) The biology of the honey bee. Cambridge, Harvard Univerty Press. 276 p. Google Scholar
39.Witherell, P.C. (1971) Duration of flight and interflight time of drone honey bees, Apis mellifera. Ann. Entomol. Soc. Am. 64(3), 609–612. Article Google Scholar
40.Woyke, J., Wilde, J., Wilde, M. (2001) Apis dorsata drone flights, collection of semen from everted endophalli and instrumental insemination of queens. Apidologie 32, 407–416. Article Google Scholar
Добавить комментарий