
Резюме
Недавние дебаты поставили под сомнение, действительно ли социальное обучение животных заслуживает ярлыка «социальное» [1]. Одиночные животные иногда могут учиться у сородичей [2, 3], а способности к социальному обучению часто коррелируют с индивидуальными способностями к обучению [4, 5, 6], поэтому может быть мало оснований рассматривать лежащие в основе обучения процессы как адаптивно специализированные. Здесь мы демонстрируем, как обучение путем наблюдения — способность, общая для приматов, птиц, грызунов и насекомых [7, 8, 9] — может возникнуть благодаря простой павловской способности интегрировать две заученные ассоциации. Известно, что шмели учатся распознавать цвета цветов, наблюдая за сородичами из-за ширмы [9], и мы обнаружили, что предыдущие ассоциации между сородичами и вознаграждением имеют решающее значение для этого процесса. Пчелы, которые ранее получали вознаграждение за присоединение к сородичам, копируют цветовые предпочтения, а пчелы, не имеющие такого опыта, нет, и те, которые ассоциируют сородичей с горькими веществами, активно избегают тех цветов, где видели другие. Наши выводы позволяют рассматривать сложное на первый взгляд явление социального обучения в рамках простой ассоциативной системы, которая характерна как для социальных, так и для одиночных видов.
Основные моменты
Мы проверяем, лежит ли обусловливание второго порядка в основе социального обучения пчел.
Пчелы, ассоциирующие сородичей с аппетитными стимулами, копируют их, а наивные пчелы — нет.
Пчелы, ассоциирующие сородичей с неприятными стимулами, избегают популярных цветов.
Любое животное, которое может интегрировать заученные ассоциации, потенциально может обучаться социально.
Результаты и обсуждение
Обусловление второго порядка — это двухэтапный процесс ассоциативного обучения, который был впервые продемонстрирован Павловым [10], который обучил собак ассоциировать звук метронома (условный стимул, CS1) с предъявлением пищи (необусловленный стимул, US). Затем он сопряг показ черного квадрата (CS2) с метрономом (CS1) в отсутствие пищи, обнаружив, что впоследствии при виде только черного квадрата (CS2) у собак начиналось слюноотделение. Таким образом, CS2 стал ассоциироваться с US, несмотря на то, что они никогда не испытывались вместе (рис. 1A). С тех пор обусловливание второго порядка было продемонстрировано у широкого круга видов, включая людей, крыс, рыб, пчел, плодовых мушек и морских слизней [11, 12, 13, 14, 15, 16, 17, 18, 19]. Его экологическая значимость редко рассматривалась, но мы ранее предположили, что он может дать мощное объяснение некоторым формам наблюдательного обучения ([20]; см. также [21, 22]).

(B) Предполагается, что тот же механизм лежит в основе наблюдательного обучения у шмелей.
Известно, что шмели запоминают цветовые предпочтения фуражиров, за которыми им было позволено наблюдать через перпексовый экран, и «копируют» эти предпочтения при самостоятельном фуражировании [9]. Здесь мы проверили, является ли такое поведение результатом обусловливания второго порядка и, следовательно, зависит ли оно от ассоциации первого порядка между визуальными сигналами конспецифических видов и пищевым вознаграждением. Согласно нашей гипотезе, пчелы копируют предпочтения демонстраторов, поскольку их кормовой опыт привел их к ассоциации визуального облика конспецификов (CS1) с пищей (US) [23]; конспецифики склонны задерживаться только на соцветиях, приносящих вознаграждение. Когда конспецификов (CS1) впоследствии видят на определенном цвете цветка (CS2), пчелы-наблюдатели формируют вторичную ассоциацию между цветом цветка (CS2) и пищевой наградой (US; рис. 1B). Два прямых предсказания заключаются в том, что пчелы, не имеющие такого опыта, не должны копировать и что пчелы, ассоциирующие сородичей с альтернативным, горьким веществом, должны активно избегать тех цветов, на которых были замечены другие пчелы.
Мы обучили отдельных пчел (n = 158; 11 колоний), не имевших опыта кормления, посещать вертикальную решетку из шести небольших прозрачных кормовых платформ на летной арене. Три платформы были заняты мертвыми или модельными родственными «демонстраторами» (по три на платформу; всего девять), а остальные три платформы были незаняты. Для испытуемых из группы «сахарозы» демонстраторов можно было найти только на платформах, содержащих раствор сахарозы; незанятые платформы содержали аверсивный раствор хинина [24]. Для пчел из группы «хинин» занятые платформы содержали раствор хинина, а незанятые альтернативы — раствор сахарозы. Пчелы в обеих группах были успешны в обучении этой задаче первого порядка (см. [23]), выбирая вознагражденную альтернативу значительно чаще, чем ожидалось случайно к заключительной тренировочной схватке (случайные ожидания: 50%; средние показатели ± SD: группа хинина, 93 ± 10%; группа сахарозы, 89 ± 13%; одновыборочный t-тест: t = 4,01, df = 49, p < 0,001. и t = 3,57, df = 52, p < 0,001, соответственно). Третья, «наивная» группа проходила такое же обучение, добывая корм с трех платформ, содержащих раствор сахарозы, и трех, содержащих раствор хинина, но в отсутствие демонстраторов. Сразу же после этой фазы обучения первого порядка мы позволили каждой пчеле в течение 10 минут наблюдать за кормовой ареной из-за экрана из персекса. Внутри арены испытуемый мог видеть массив из шести цветных цветков двух типов — либо три оранжевых и три зеленых, либо три синих и три желтых, все на коричневом фоне. В каждом случае все цветы одной из цветовых альтернатив были заняты демонстраторами. После этого периода наблюдения мы меняли цветы на чистые, незанятые, пустые заменители тех же цветов и изменяли пространственное расположение цветов. Затем испытуемого выпускали на арену и регистрировали количество приземлений на каждый цвет цветка. Как и ожидалось в соответствии с нашей гипотезой, поведение копирования отражало предшествующие ассоциации, которые пчелы развили между родственниками и аверсивными или аппетитными необусловленными стимулами (рис. 2). Пчелы, которые ассоциировали конспецификов с сахарозой, с большей вероятностью посещали цветки, если видели там демонстраторов, чем если демонстраторы были замечены на альтернативном цвете (F = 5,27, df = 1, p = 0,02); обратное было верно для пчел, которые ассоциировали конспецификов с хинином (F = 4,23, df = 1, p = 0,04). Пчелы в наивной группе не показали значительного предпочтения ни в одном из направлений (F = 1,21, df = 1, p = 0,28). Данные по тестам выбора оранжевого/зеленого и синего/желтого были объединены, так как мы не обнаружили значительного влияния теста выбора на данные (F < 1,5, df = 1, p > 0,2 в каждом случае).
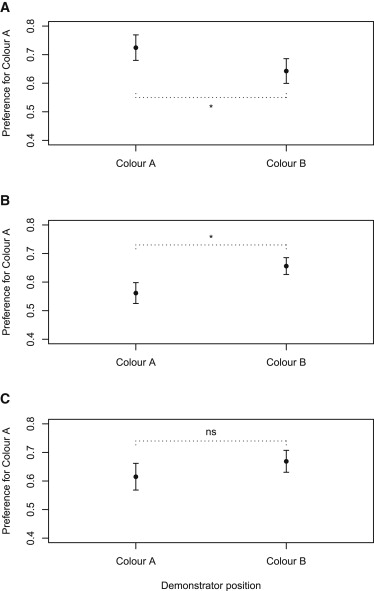
Социальное изучение пищи обычно включает в себя привлечение к местам кормления или типам пищи. Однако предыдущие авторы отмечали, что если использование социальной информации является формой ассоциативного обучения, то животные должны быть способны усваивать негативные ассоциации так же, как и позитивные [1, 25, 26]. Другими словами, если присутствие сородичей обратно коррелирует с качеством или доступностью пищи, животные должны избегать популярных ресурсов. Наши данные представляют собой эмпирическое доказательство того, что отрицательные связи действительно могут приводить к избегающему поведению. Более того, нет ничего нереалистичного в том, чтобы ожидать, что дикие кормовые пчелы могут иногда научиться отрицательно ассоциировать сородичей с пищей [27], поскольку сородичи уничтожают цветы. Мы экстраполируем наш вывод с осторожностью, потому что активный аверсивный стимул, такой как хинин, может быть более эффективным в вызывании поведения избегания, чем просто отсутствие пищи. Тем не менее, возможность того, что дикие пчелы, скорее всего, будут избегать видов цветов, где были замечены сородичи, а не предпочитать их, заслуживает дальнейшего изучения.
В традиционной терминологии социального обучения копирование цвета у шмелей можно назвать «усилением стимула» [28], когда присутствие пчел-демонстраторов знакомит наблюдателя с цветом цветка, возможно, привлекая к нему внимание. В противном случае, если присутствие демонстратора вызывает у наблюдателей ассоциацию между цветом цветка и вознаграждением, можно использовать термин «наблюдательное обусловливание» [7]. Хотя оба механизма могут быть включены в рамки индивидуального обучения (обучение с одним стимулом или обучение со стимулом, соответственно [24]), эти объяснения не затрагивают вопрос о том, почему поведение демонстратора приводит к такому воздействию. Предполагается, что отбор привел к тому, что социальные животные рассматривают поведение конспецификов как биологически важный стимул. Обусловление второго порядка предполагает отсутствие такой запрограммированной реакции на социальные сигналы [24].
Наши результаты не означают, что копирование цветовых предпочтений у шмелей не связано с адаптивной специализацией. Особь может с большей вероятностью научиться ассоциациям первого порядка с участием сородичей, если она имеет зрительные системы, приспособленные для восприятия социальных сигналов, перцептивные системы, которые отдают им приоритет, или мотивационные системы, которые поощряют близость [1, 29, 30], хотя существует мало доказательств того, что это так у шмелей [31]. Другими словами, механизмы ввода (ср. «механизмы сбора данных» [32]) вполне могут быть приспособлены для преимущественной обработки социальной информации [1]. Новизна наших результатов заключается скорее в том, что, хотя отбор может благоприятствовать животным, которые отдают предпочтение социальным стимулам, ассоциативное обучение может объяснить, почему эти стимулы приобретают актуальность. Таким образом, любое животное, способное к обусловливанию второго порядка, должно быть способно и к этой форме наблюдательного обучения при соответствующих обстоятельствах.
Мы показали, что обучение путем наблюдения может развиваться, если животные учатся ассоциировать пищу, биологически важный стимул, с визуальным присутствием сородичей. Когда эти визуальные сигналы затем наблюдаются в контексте другого биологического стимула, этот второй стимул также ассоциируется с пищей посредством простого павловского процесса. Важно отметить, что нет никаких причин для того, чтобы условные стимулы первого порядка генерировались исключительно конспецификами. Если гетероспецифическое поведение может быть использовано для предсказания вознаграждения, то обусловливание второго порядка дает животным возможность воспользоваться такой информацией, и появляется все больше доказательств того, что социальное обучение действительно преодолевает видовые границы [31, 33, 34, 35, 36].
Экспериментальные процедуры
Подопытные животные и арена
Все экспериментальные процедуры были одобрены Комитетом по этике Лондонского зоологического общества. Одиннадцать колоний шмелей (Bombus terrestris) (любезно предоставленных Syngenta Bioline Bees, Weert, Нидерланды) были размещены в деревянных гнездовых ящиках (28 × 16 × 11 см), соединенных плексигласовой трубой с летной ареной (117 × 72 × 30 см). Перед началом эксперимента пчел кормили через гравитационные кормушки внутри гнездового ящика и индивидуально помечали пронумерованными бирками. Поскольку наши шмелиные гнездовья содержатся в темноте, у особей не было возможности ассоциировать визуальное присутствие сородичей с пищей.
Первая фаза: обучение
Мы обучили более 250 пчел, и 158 из них завершили тест без потери мотивации к добыче корма (n = 54, 50 и 54 в группах с сахарозой, наивных и хинином, соответственно). Испытуемые обучались в одиночку. Каждому из них разрешалось посетить массив из шести кормушек с прозрачными платформами (1,5 × 1 × 1,5 см; два вертикальных ряда по три платформы в каждом, установленные на картонном фоне). Девять демонстраторов (см. ниже) были установлены вокруг трех платформ (по три демонстратора на платформу, два приколоты и один болтается на нитке, чтобы обеспечить движение при включении вентилятора); остальные три были незаняты.
В качестве демонстраторов выступали модельные пчелы или мертвые неродственные рабочие пчелы, убитые замораживанием в день, предшествующий эксперименту. Модели были сделаны из запекаемой в духовке глины (Fimo soft, Staedtler) и окрашены так, чтобы отразить естественные цветовые свойства Bombus terrestris в соответствии с цветовым пространством шмелей [37, 38]. Мы использовали модели, чтобы не убивать чрезмерное количество пчел; в предыдущей работе было обнаружено, что модели, мертвые пчелы и живые пчелы вызывают эквивалентное поведение копирования [9, 39]. Модели не издают запаха, но на этапе тестирования (см. ниже) мы наблюдали за ними из-за экрана, поэтому в обоих случаях были доступны только визуальные сигналы.
В группе с сахарозой три занятые платформы содержали 20 мкл 2 М раствора сахарозы, а незанятые платформы содержали 20 мкл насыщенного раствора гемисульфата хинина. В группе с хинином занятые платформы содержали хинин, а незанятые — раствор сахарозы. Пчелы в наивной группе посещали тот же массив, где три платформы содержали раствор сахарозы, а три — раствор хинина, но без демонстраторов. Небольшой ручной вентилятор («The Cooler» от Design Go; 8,5 × 4,5 × 4 см) был включен во время фазы обучения, чтобы создать видимость движения пчел-демонстраторов (в соответствии с [9]).
Когда пчелы собирают полный урожай раствора сахарозы, они кладут его в гнездо, а затем снова возвращаются за кормом. Во время фазы обучения подопытной пчеле разрешалось возвращаться восемь раз; каждый раз положение занятых и незанятых платформ менялось, чтобы избежать того, что испытуемые заучивали только положение цветков с вознаграждением. Платформы пополнялись каждый раз, когда они истощались, и регистрировалось количество посадок на занятые и незанятые платформы.
Вторая фаза: наблюдение
Наша фаза наблюдения была разработана по аналогии с фазой, использованной Ворденом и Папаем [9]. По возвращении на арену после внесения последнего груза сахарозы, собранного во время обучения, каждый испытуемый помещался в наблюдательный бокс, прикрепленный к боковой стенке арены и содержащий ультрафиолетовый прозрачный перспексный экран (21 × 22 см), через который можно было видеть кормовую решетку с расстояния 11 см. Учебная кормовая площадка была заменена на аналогичную, где каждая кормовая площадка была отмечена цветным картонным «цветком» (диаметром 57 мм) на коричневом картонном фоне. Шесть цветков состояли либо из трех оранжевых и трех зеленых цветков, либо из трех синих и трех желтых цветков. В рамках набора цветов для выбора, цвета были случайным образом распределены по платформам. Цветовой контраст между каждым цветом и фоном, рассчитанный в цветовом пространстве для пчел [37, 38], был немного выше для синего (0,21), чем для желтого (0,18), и умеренно выше для оранжевого (0,2), чем для зеленого (0,14), где максимальный теоретический контраст между любым цветом и фоном равен 1. Каждая кормовая платформа была заполнена 20 мкл воды.
Три пчелы-демонстратора были прикреплены к цветкам одного цвета в тех же положениях, которые были описаны в настройках фазы обучения. Для перемещения подвешенных демонстраторов снова использовался вентилятор. В течение 10 минут испытуемым разрешалось наблюдать за цветочным массивом с демонстраторами. Вертикальное расположение массива означало, что все цветы были одинаково видны из-за экрана. В середине периода наблюдения положение оранжевого и зеленого или синего и желтого цветов менялось местами, чтобы минимизировать вероятность того, что пчелы ассоциируют демонстраторы с местоположением, а не с цветом.
Третья фаза: Тестирование
Сразу после периода наблюдения цветочный массив был заменен идентичным цветочным массивом, за исключением того, что расположение каждого цвета цветка было случайным образом перераспределено, и, что важно, демонстраторы не присутствовали. Все платформы для кормления были заполнены 20 мкл воды. Испытуемого выпускали на арену, и в течение 5 минут после (и включая) первого приземления на цветок регистрировали его выбор посадки. Если в течение 10 минут после выпуска испытуемого не было совершено ни одного приземления, тест прекращался.
Тест на предпочтение цвета
Наша экспериментальная схема включала два типа теста на выбор: оранжевый/зеленый и синий/желтый, поэтому переменными ответа были «предпочтение оранжевого» или «предпочтение синего» соответственно. Чтобы сравнить врожденные предпочтения этих цветов, мы протестировали 30 абсолютно наивных пчел, которые никогда ранее не видели цветных стимулов, на каждой паре цветов. Оранжевый цвет умеренно предпочтительнее зеленого, а синий — желтого (среднее ± SE: 0,64 ± 0,04 и 0,60 ± 0,05, соответственно), как и следовало ожидать, исходя из различий в цветовом контрасте с фоном (см. раздел «Вторая фаза: наблюдение» выше). Эти предпочтения для оранжевого и синего цветов существенно не отличаются друг от друга (F < 0,01, df = 1, p = 0,95), и, соответственно, данные были объединены для двух тестов выбора, а «тест выбора» был включен в качестве предиктора в каждую модель.
Статистические тесты
Наша гипотеза предсказывает, что предпочтение любого цвета цветка должно быть относительно больше, когда демонстраторы, связанные с сахарозой, видны на этом цвете, чем когда они видны на альтернативном выборе. Обратное предсказывается для демонстраторов, связанных с хинином, тогда как пчелы в наивной группе должны быть безразличны к присутствию демонстраторов. Таким образом, для каждой из трех групп обработки мы смоделировали переменную ответа «предпочтение цвета А», где цвет А был оранжевым или синим, соответственно. В качестве «цвета А» мы выбрали оранжевый/синий, так как врожденные предпочтения этих двух цветов существенно не различаются (см. раздел «Тест цветовых предпочтений» выше). Обратите внимание, что предпочтения к цвету B прямо противоположны предпочтениям к цвету A, поэтому моделирование этих предпочтений даст идентичные результаты.
Для каждой группы мы смоделировали долю посещений цветов каждым испытуемым, направленных на цвет А, используя обобщенные линейные модели с квазибиномиальной структурой ошибок [39]. Положение демонстраторов (цвет A или цвет B), тест выбора (оранжевый/зеленый или синий/желтый) и доля правильных выборов в финальной тренировочной схватке были подобраны в качестве предикторов. Незначимые члены последовательно исключались до тех пор, пока дальнейшее упрощение существенно не снижало объяснительную способность модели. Значения p представляют эффект удаления значимых членов из минимальной модели, оцененный с помощью F-тестов [40].
Благодарности
Э.Л. финансировалась стипендией ранней карьеры от Leverhulme Trust. А.А.-В. был профинансирован постдокторской стипендией Фонда Фиссена. Мы благодарим Шарлотту Локвуд и Адама Девениша за помощь в сборе данных и трех анонимных рецензентов за комментарии к рукописи.
Ссылки
1.Heyes C.
What’s social about social learning? .J. Comp. Psychol. 2012; 126: 193-202
Scopus (252) PubMed Crossref Google Scholar
2. Wilkinson A. Kuenstner K. Mueller J.Huber L.
Social learning in a non-social reptile (Geochelone carbonaria).Biol. Lett. 2010; 6: 614-616
Scopus (141) PubMed Crossref Google Scholar
3. Fiorito G. Scotto P.
Observational-learning in Octopus vulgaris.Science. 1992; 256: 545-547
Scopus (265) PubMed Crossref Google Scholar
4. Lefebvre L. Giraldeau L.A.
Is social learning an adaptive specialization?.in: Heyes C.M. Galef B.G. Social Learning in Animals: The Roots of Culture. Academic Press, Inc., San Diego, CA, USA1996
Google Scholar
5. Reader S.M. Hager Y. Laland K.N.
The evolution of primate general and cultural intelligence.Philos. Trans. R. Soc. Lond. B Biol. Sci. 2011; 366: 1017-1027
Scopus (287) PubMed Crossref Google Scholar
6. Bouchard J. Goodyer W. Lefebvre L.
Social learning and innovation are positively correlated in pigeons (Columba livia).Anim. Cogn. 2007; 10: 259-266
Scopus (110) PubMed Crossref Google Scholar
7. Cook M. Mineka S.
Observational conditioning of fear to fear-relevant versus fear-irrelevant stimuli in rhesus monkeys.J. Abnorm. Psychol. 1989; 98: 448-459
Scopus (218) PubMed Crossref Google Scholar
8. Galef Jr., B.G. Giraldeau L.A.
Social influences on foraging in vertebrates: causal mechanisms and adaptive functions.Anim. Behav. 2001; 61: 3-15
Scopus (514) PubMed Crossref Google Scholar
9. Worden B.D. Papaj D.R.
Flower choice copying in bumblebees.Biol. Lett. 2005; 1: 504-507
Scopus (116) PubMed Crossref Google Scholar
10. Pavlov, I.P. (1927).
Conditioned Reflexes: An Investigation of the Physiological Activity of the Cerebral Cortex, G.V. Anrep, trans. and ed. (Mineola, NY, USA: Dover).
Google Scholar
11. Amiro T.W. Bitterman M.E.
Second-order appetitive conditioning in goldfish.J. Exp. Psychol. Anim. Behav. Process. 1980; 6: 41-48
Scopus (20) PubMed Crossref Google Scholar
12. Holland P.C. Rescorla R.A.
Second-order conditioning with food unconditioned stimulus.J. Comp. Physiol. Psychol. 1975; 88: 459-467
Scopus (146) PubMed Crossref Google Scholar
13. Hussaini S.A. Komischke B. Menzel R. Lachnit H.
Forward and backward second-order Pavlovian conditioning in honeybees.Learn. Mem. 2007; 14: 678-683
Scopus (32) PubMed Crossref Google Scholar
14. Jara E. Vila J. Maldonado A.
Second-order conditioning of human causal learning.Learn. Motiv. 2006; 37: 230-246
Scopus (16) Crossref Google Scholar
15. Hawkins R.D. Greene W. Kandel E.R.
Classical conditioning, differential conditioning, and second-order conditioning of the Aplysia gill-withdrawal reflex in a simplified mantle organ preparation.Behav. Neurosci. 1998; 112: 636-645
Scopus (34) PubMed Crossref Google Scholar
16. Tabone C.J. de Belle J.S.
Second-order conditioning in Drosophila.Learn. Mem. 2011; 18: 250-253
Scopus (26) PubMed Crossref Google Scholar
17. Takeda K.
Classical conditioned response in the honey bee.J. Insect Physiol. 1961; 6: 168-179
Scopus (276) Crossref Google Scholar
18. Grossmann K.E.
Belohnungsverzögerung beim Erlernen einer Farbe an einer künstlichen Futterstelle durch Honigbienen.Z. Tierpsychol. 1971; 29: 28-41
Scopus (23) Crossref Google Scholar
19. Bitterman M.E. Menzel R. Fietz A. Schäfer S.
Classical conditioning of proboscis extension in honeybees (Apis mellifera).J. Comp. Psychol. 1983; 97: 107-119
Scopus (834) PubMed Crossref Google Scholar
20. Leadbeater E. Chittka L.
Social learning in insects—from miniature brains to consensus building.Curr. Biol. 2007; 17: R703-R713
Scopus (242) PubMed Abstract Full Text Full Text PDF Google Scholar
21. Giurfa M.
Social learning in insects: a higher-order capacity?.Front Behav. Neurosci. 2012; 6: 57
PubMed Google Scholar
22. Avarguès-Weber A. Deisig N. Giurfa M.
Visual cognition in social insects.Annu. Rev. Entomol. 2011; 56: 423-443
Scopus (124) PubMed Crossref Google Scholar
23. Leadbeater E. Chittka L.
Bumble-bees learn the value of social cues through experience.Biol. Lett. 2009; 5: 310-312
Scopus (63) PubMed Crossref Google Scholar
24. Chittka L. Dyer A.G. Bock F. Dornhaus A.
Psychophysics: bees trade off foraging speed for accuracy.Nature. 2003; 424: 38
Scopus (296) PubMed Crossref Google Scholar
25. Heyes C.M.
Social learning in animals: categories and mechanisms.Biol. Rev. Camb. Philos. Soc. 1994; 69: 207-231
PubMed Crossref Google Scholar
26. Hoppitt W. Laland K.
Social processes influencing learning in animals: A review of the evidence.Adv. Study Behav. 2008; 38: 105-165
Scopus (208) Crossref Google Scholar
27. Kawaguchi L.G. Ohashi K. Toquenaga Y.
Contrasting responses of bumble bees to feeding conspecifics on their familiar and unfamiliar flowers.Proc. Biol. Sci. 2007; 274: 2661-2667
Scopus (54) PubMed Crossref Google Scholar
28. Spence K.W.
Experimental studies of learning and the higher mental processes in infra-human primates.Psychol. Bull. 1937; 34: 806-85
Scopus (125) Crossref Google Scholar
29. Shettleworth S.J. Cognition,
Evolution and Behaviour. Oxford University Press, New York1998
Google Scholar
30. Marler P. Slabbekoorn H.
Nature’s Music: The Science of Birdsong. Elsevier Academic Press, San Diego, CA, USA2004
Google Scholar
31. Dawson E.H. Chittka L.
Conspecific and heterospecific information use in bumblebees.PLoS ONE. 2012; 7: e31444
Scopus (57) PubMed Crossref Google Scholar
32. Lotem A. Halpern J.Y.
Coevolution of learning and data-acquisition mechanisms: a model for cognitive evolution.Philos. Trans. R. Soc. Lond. B Biol. Sci. 2012; 367: 2686-2694
Scopus (46) PubMed Crossref Google Scholar
33. Goodale E. Nieh J.C.
Public use of olfactory information associated with predation in two species of social bees.Anim. Behav. 2012; 84: 919-924
Scopus (27) Crossref Google Scholar
34. Seppänen J.T. Forsman J.T.
Interspecific social learning: novel preference can be acquired from a competing species.Curr. Biol. 2007; 17: 1248-1252
Scopus (89) PubMed Abstract Full Text Full Text PDF Google Scholar
35. Goodale E. Beauchamp G. Magrath R.D. Nieh J.C. Ruxton G.D.
Interspecific information transfer influences animal community structure.Trends Ecol. Evol. 2010; 25: 354-361
Scopus (250) PubMed Abstract Full Text Full Text PDF Google Scholar
36. Avarguès-Weber A. Dawson E.H. Chittka L.
Mechanisms of social learning across species boundaries.J. Zool. 2013; (Published online January 17, 2013)https://doi.org/10.1111/jzo.12015
Scopus (43) Crossref Google Scholar
37. Chittka L.
The colour hexagon- A chromaticity diagram based on photoreceptor excitations as a generalized representation of color opponency.J. Comp. Physiol. A. 1992; 170: 533-543
Google Scholar
38. Stelzer R.J. Raine N.E. Schmitt K.D. Chittka L.
Effects of aposematic coloration on predation risk in bumblebees? A comparison between differently coloured populations, with consideration of the ultraviolet.J. Zool. 2010; 282: 75-83
Scopus (16) Crossref Google Scholar
39. Leadbeater E. Chittka L.
The dynamics of social learning in an insect model, the bumblebee (Bombus terrestris).Behav. Ecol. Sociobiol. 2007; 61: 1789-1796
Scopus (101) Crossref Google Scholar
40. Crawley M.J.
The R Book.John Wiley & Sons, Ltd., Chichester, UK2007
Scopus (5165) Crossref Google Scholar
Добавить комментарий