
Аннотация
Одичавшие колонии Apis mellifera широко распространены по всему миру и оказывают экологическое воздействие как опылители и конкуренты за пищу и возможность гнездования. Величина воздействия зависит от плотности их популяции, но знания об этой плотности недостаточны. Мы зафиксировали 69 колоний одичавших A. mellifera на км2 во фрагментированных эвкалиптовых лесах в Австралии, что превышает оценки, полученные в других странах мира, и сравнимо только с одним другим австралийским исследованием. Мы обследовали 52,5 га лесных участков с 357 гнездовыми ящиками, установленными для обеспечения возможности гнездования находящихся под угрозой исчезновения позвоночных животных. Наши участки охватывали регион более 140 км в поперечнике с повторными исследованиями в течение 3-6 лет. Мы показали, что на использование гнездовых ящиков одичавшими колониями A. mellifera влияет дизайн ящика (p = 0,042), со слабыми доказательствами интерактивного эффекта типа растительности на участке (остатки леса против подсадки) и древесного покрова в пределах 500 м (p = 0,091). При плотности 69 колоний на км2 эта плотность эквивалентна рекомендуемому количеству ульев для опыления некоторых сельскохозяйственных культур и, следовательно, может влиять на опыление и приводить к конкуренции с другими посетителями цветка. Apis mellifera также может конкурировать за дупла с местной фауной, зависящей от полостей, особенно в ландшафтах, где были удалены деревья.
Введение
Западная медоносная пчела (Apis mellifera) широко используется по всему миру, вызывая значительные экологические последствия как опылитель и как конкурент за гнездовые и пищевые ресурсы (1). Apis mellifera также избежала одомашнивания, создав одичавшие популяции во многих странах (2,3,4), где она может оказывать негативное воздействие на местное биоразнообразие (5,6,7,8). Эти одичавшие пчелы также играют важную роль в опылении экономически значимых культур (9) и местных растений (4). Считается, что численность одичавших колоний A. mellifera в дикой природе сократилась во многих регионах мира, особенно по мере распространения паразитического клеща Varroa destructor из Азии в остальной мир (10,11). В то же время африканизированные медоносные пчелы (т.е. пчелы европейского происхождения, которые гибридизировались с подвидом A. m. scutellata) распространились в Америке и показали различные уровни толерантности к Varroa (12), так что новая волна инвазии привела к новым последствиям для сетей опылителей и результатов опыления (13,14). Однако пространственное распределение одичавших колоний A. mellifera плохо изучено из-за сложности сбора данных.
Как эусоциальное насекомое, западная медоносная пчела была признана особенно влиятельным инвазивным видом, поскольку она может быстро увеличивать численность популяции в своем инвазивном ареале и использовать ресурсы (такие как нектар и пыльца) с помощью скоординированной когорты фуражиров, что превышает возможности несоциальных насекомых в том же регионе (15). Хотя можно обоснованно ожидать, что экологическое воздействие одичавшей A. mellifera будет зависеть от плотности колонии (16) , не существует установленного базового уровня для определения того, когда можно ожидать возникновения воздействия, имеющего значение для управления охраной природы. Тем не менее, мы можем сравнить рекомендуемую «норму заселения ульев» для управляемых ульев, используемых для опыления сельскохозяйственных культур, с плотностью одичавших колоний. Плотность, достаточно высокая для обеспечения опыления монокультурной культуры, может считаться высокой с точки зрения взаимодействия пчел с цветами местной растительности. Нормы заселения ульев сильно варьируются в зависимости от типа культур, но недавний обзор показал, что рекомендуемые нормы варьируются от примерно 50 до более чем 1000 колоний на км2 (17).
Для размещения своих колоний одичавшие пчелы A. mellifera в основном используют дупла деревьев, хотя при наличии возможностей также используются искусственные сооружения, такие как стенные полости в зданиях или коробки водомеров [например, ссылка 18]. Прямые исследования одичавших колоний являются трудоемкими, поскольку этот вид может жить в дуплах, расположенных высоко над землей и широко разбросанных по ландшафту. В результате, лишь немногие исследования сообщают о закономерностях, полученных при обследовании больших территорий. Ратниекс и др. (19) собрали данные девяти исследований по всему миру и сообщили о плотности от 0,17 до 9 колоний на км2. Впоследствии Баум и др. (20) зарегистрировали плотность 12,5 колоний на км2 в прибрежной прерии в Техасе, США, хотя аналогичное исследование, проведенное 13 лет спустя, показало, что плотность снизилась до 5,4 колоний на км2 (21). Олдройд и др. (22) исследовали 35 га леса в Виктории, Австралия, и сообщили о плотности, которая в разные годы варьировалась от 50 до 150 колоний на км2. Плотность, о которой сообщалось в работе Oldroyd et al. (22), была названа самой высокой из когда-либо зарегистрированных, что ставит ее в диапазон норм заселения ульев для опыления сельскохозяйственных культур, и поэтому она была охарактеризована как нетипичная (23). Однако, учитывая нехватку подобных исследований во всем мире, основа для сравнения между исследованиями невелика.
Как и другие виды фауны, зависящие от дупла, их зависимость от дупла деревьев означает, что одичавшие популяции A. mellifera могут быть сокращены в результате вырубки деревьев, а последующее восстановление может быть медленным, поскольку развитие дупла происходит медленно и, как правило, на старых деревьях (24). При наличии нескольких видов, конкурирующих за ограниченные полости деревьев, также возможно, что успешные захватчики, такие как A. mellifera, могут вытеснить местную фауну (25). По этой причине конкуренция с одичавшими медоносными пчелами включена в список основных угрожающих процессов для исчезающих видов деревьев, обитающих в полостях, в законодательстве об охране окружающей среды австралийского штата Новый Южный Уэльс (26). Одной из стратегий, обычно применяемых для восстановления популяций сокращающейся фауны, зависящей от дупла, является установка искусственных гнезд (27) , но если эти гнезда заняты медоносными пчелами, то эта стратегия может быть подорвана. Сообщается, что медоносные пчелы пользуются гнездовыми ящиками во многих ландшафтах (28,29,30), а в некоторых случаях они являются одними из самых частых пользователей этих искусственных сооружений (31,32).
Оценка использования искусственных гнездовых ящиков A. mellifera более доступна, чем исследование естественных дуплах, поскольку местоположение гнездовых ящиков известно и может контролироваться. Однако в исследованиях искусственных гнездовых ящиков обычно сообщается только о доле занятых гнездовых ящиков [например, ссылка 31]. Хотя это дает представление о потенциальной конкуренции за возможность гнездования, это не позволяет рассчитать ландшафтную плотность колоний медоносных пчел, что особенно важно для понимания воздействия этого вида на экосистему.
Другой метод, который развивается для оценки плотности колоний A. mellifera в больших масштабах, заключается в отлове самцов (трутней) с феромонной приманкой и последующем использовании генетических методов для различения различных братских групп (33,34). Этот подход требует подтверждения ряда предположений, включая расстояние полета трутней и долю колоний, вносящих трутней в выборку. Утайпанон и др. (35) показали, что дроны могут пролететь до 3,75 км до приманки, но данных было недостаточно (только два трансекта) для оценки доверительных интервалов вокруг этой оценки. Кроме того, недавние полевые испытания показали, что только 64% известных колоний в радиусе 1,55 км внесли трутней в отловленную выборку, причем меньшее количество трутней было внесено ульями, расположенными дальше (36). Кроме того, даже при больших размерах выборки среднее количество трутней, внесенных каждой колонией, было меньше количества, необходимого для точной идентификации братских групп (36). Как следствие, расчетные плотности колоний, основанные на отлове дронов, трудно интерпретировать и, вероятно, они существенно занижают истинную плотность колоний на данной территории, что делает невозможным прямое сравнение между плотностями, установленными по данным наземных исследований, и плотностями, полученными в результате исследований с отловом дронов [например, ссылка 21].
В данном исследовании мы проанализировали использование искусственных гнездовых ящиков колониями A. mellifera в регионе южного Нового Южного Уэльса, Австралия, и задались вопросом, достаточно ли высока плотность колоний инвазивных одичавших пчел в этом ландшафте, чтобы обоснованно ожидать значительного воздействия на местное биоразнообразие. Объединив данные нескольких исследований, проведенных в одном регионе, мы собрали большой массив данных, включающий 566 гнездовых ящиков, каждый из которых обследовался не менее трех раз в течение четырех лет. Данные были уточнены до подмножества из 357 гнездовых ящиков, использованных для оценки плотности колоний, и другого подмножества из 299 гнездовых ящиков, использованных для моделирования вероятности заселения, включая структуру растительности (путем сравнения остаточных лесов и вновь посаженных лесов), лесной покров в масштабе ландшафта и дизайн гнездовых ящиков.
Использование искусственных гнездовых ящиков колониями одичавших медоносных пчел устанавливает нижнюю границу истинной плотности колоний в этом ландшафте, поскольку неучтенные колонии могут также встречаться в естественных дуплах или в других сооружениях. Поскольку мы изучали использование недавно установленных гнездовых ящиков, данное исследование дает представление о том, сильно ли возможности гнездования ограничивают плотность одичавших колоний A. mellifera на этой территории. Высокий уровень использования ящиков пчелами предполагает, что новые возможности гнездования, предоставленные искусственными гнездовыми ящиками, позволили расширить ранее существовавшую популяцию. Если использование гнездовых ящиков редкое, то это указывает либо на то, что устоявшаяся популяция пчел (т.е. источник для новых колоний) не расширяется, либо на то, что существует так много лучших возможностей для гнездования, что искусственные гнездовые ящики не так привлекательны, как доступные природные ресурсы.
Методы
Полевые участки и исследования
Искусственные гнездовые ящики были прикреплены к деревьям на 23 участках в ЮВ НЮУ и располагались в основном в пределах 10 км от шоссе Хьюм в полосе, простирающейся от городов Албери на юго-западе до Ясс на северо-востоке, с дополнительным скоплением участков в 40 км к северу от шоссе (рис. 1). Самый северный участок был отделен от самого южного участка 148 км.
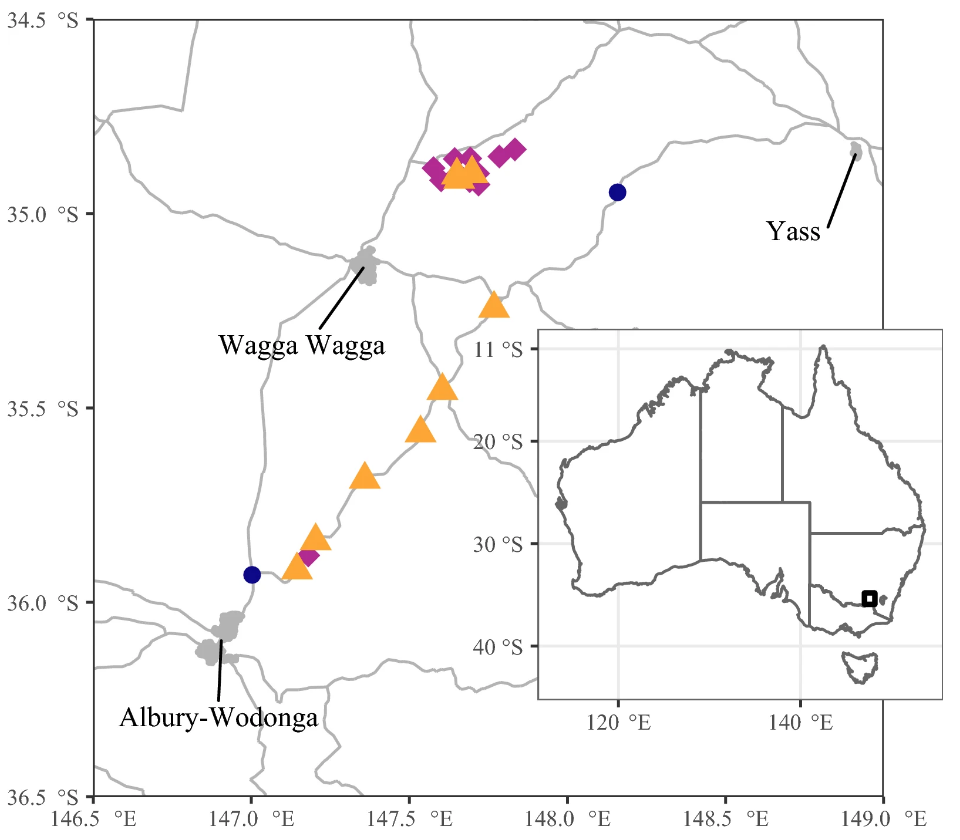
Деревянные гнездовые ящики различных конструкций были установлены с учетом требований широкого спектра различных видов фауны, использующих дупла. Гнездовые ящики находились на месте не менее восьми месяцев до первого исследования. Первые исследования на предмет занятости гнездовых ящиков были проведены в июле 2010 года, а последние — в ноябре 2014 года. Между повторными исследованиями прошло не менее четырех месяцев. На участках находилось от шести до 72 гнездовых ящиков, причем 566 ящиков в общей сложности были обследованы не менее трех и не более шести раз за период исследований (в среднем = 5,11 исследований на участок).
Во время исследований ящики визуально осматривались на предмет наличия в них людей или признаков использования. Отмечалось наличие живых колоний медоносных пчел или свидетельств того, что пчелы использовали ящик, но теперь отсутствуют, например, наличие сотов. Также были отмечены другие животные, но они не были предметом данного исследования. Работа проводилась в соответствии с разрешением на этическое отношение к животным, одобренным Австралийским национальным университетом и Управлением по охране окружающей среды и наследия Нового Южного Уэльса.
Ландшафтные метрики
Доминирующим типом растительности в этом ландшафте до расчистки под сельское хозяйство были эвкалиптовые леса умеренного пояса (37). Большинство боксов было размещено в остаточных лесных массивах, оставленных и фрагментированных, когда остальная часть ландшафта была расчищена под сельское хозяйство. Некоторые участки включали лесные массивы, которые были созданы путем пересадки ранее расчищенной территории и поэтому поддерживали более молодые деревья. Мы называли эти участки «посадками» и размещали ящики только на деревьях высотой более 6 метров. Некоторые участки были полностью состояли из пересаженных деревьев, а один большой участок представлял собой сложную смесь остатков и посаженных лесов.
Чтобы определить площадь участка, мы использовали выпуклый корпус вокруг мест расположения гнездовых ящиков для создания многоугольников и добавили 15-метровый буфер для поддержания связи в многоугольнике через разбросанные деревья леса. Этот буфер был необходим, поскольку деревья в этих лесах расположены на большом расстоянии друг от друга, даже там, где не проводилась расчистка территории. Затем мы пересекли полигоны с картой полога деревьев с разрешением 5 м, основанной на снимках SPOT5 за период 2008-2011 (38) гг. Этот анализ был выполнен в программе R (39).
Некоторые участки находились в придорожных полосах растительности и поэтому были длинными и узкими, что затрудняло расчет значимой площади участка. С целью вычисления плотности пчелиных колоний мы сократили набор данных до тех участков, площадь которых превышала 1 га, и исключили одиннадцать самых маленьких и узких участков. Таким образом, осталось десять участков площадью от 2,4 до 12,8 га, с общей площадью 52,5 га, содержащих 357 гнездовых ящиков.
Для каждого гнездового ящика мы также рассчитали количество древесного покрова в близлежащем ландшафте, используя тот же слой древесного покрова38. Мы рассчитали долю окружности радиусом 100 м с центром на каждом гнездовом ящике, которая была занята древесным покровом, а затем рассчитали ту же долю в окружности радиусом 500 м. Поскольку в центре всегда находилось дерево (т.е. дерево с гнездовым ящиком), рассчитанные пропорции всегда были больше нуля.
Статистическое моделирование
В ходе всего исследования было установлено 16 различных конструкций ящиков. Для статистического моделирования мы уточнили набор данных, чтобы исключить типы гнездовых ящиков, для которых было менее 25 повторов. Мы также исключили один большой участок, на котором перемежались остатки и посаженные деревья, поскольку такая структура была нетипичной для нашего проекта и для более широкого ландшафта. Уточненный набор данных включал 21 участок с 299 гнездовыми ящиками четырех различных типов (табл. 1, 2). Пять участков включали остатки и посадки, десять — только остатки, а шесть участков полностью состояли из пересаженных деревьев (Таблица 2). Типы гнездовых ящиков включали ящики среднего размера для небольших планеров и фаскогалов (Petaurus breviceps, Phascogale tapoatafa, тип A), более крупные ящики, предназначенные для дендроидных млекопитающих, таких как беличьи планеры (Petaurus norfolcensus, тип B), ящики среднего размера для попугаев (включая превосходного попугая, Polytelis swainsonii, тип C) и меньшие ящики для птиц, таких как бурый древолаз (Climacteris picumnus, тип D).
| Тип | Глубина (мм) | Ширина (мм) | Высота (мм) | Объем (литры) | Диаметр входа (мм) | Площадь входа (мм2) |
| А | 170 | 170 | 500 | 14.5 | 40 | 1257 |
| B | 300 | 300 | 500 | 45 | 80 | 5027 |
| C | 200 | 200 | 550 | 22 | 90 | 6362 |
| D | 150 | 150 | 150 | 3.4 | 50 | 1964 |
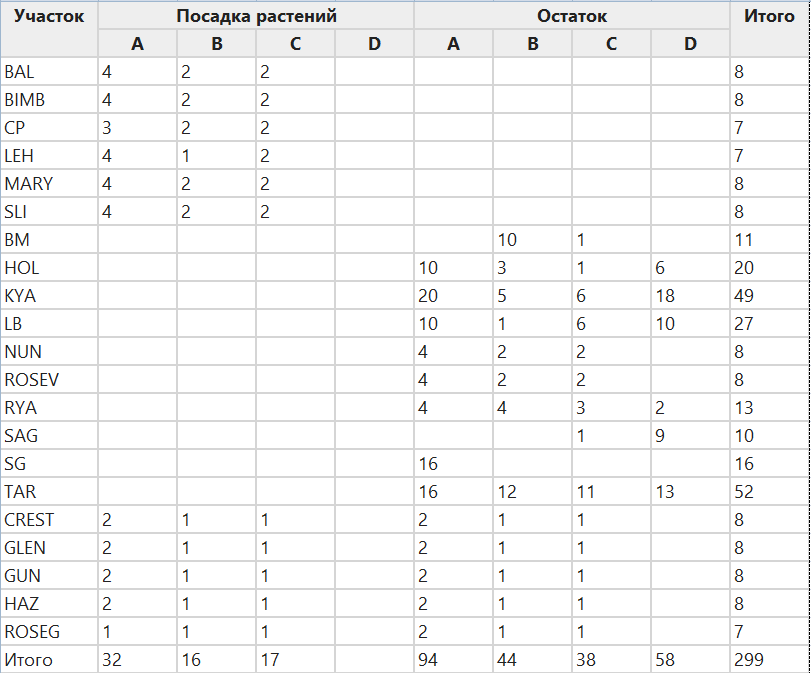
Чтобы проверить, какие факторы предсказывают наличие колоний медоносных пчел в гнездовых ящиках, мы использовали обобщенное линейное моделирование. Переменной отклика было наличие или отсутствие колоний пчел в гнездовых ящиках весной 2011 года. Нашими переменными-предикторами были тип гнездового ящика, тип участка, доля древесного покрова в пределах 100 м от участка, доля древесного покрова в пределах 500 м от участка и участок (Таблица 3).
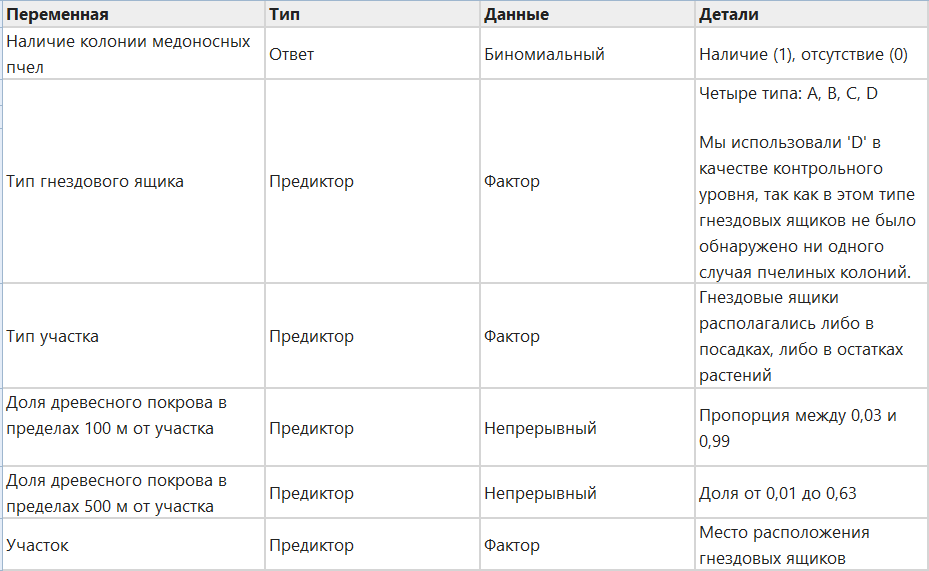
Мы использовали процедуру усреднения модели с использованием информационного критерия Акаике с поправкой на малые размеры выборки (AICc 40). Вначале мы установили обобщенную линейную смешанную модель (GLMM), содержащую все предикторные переменные и их взаимодействия с переменной присутствия/отсутствия колоний медоносных пчел, предполагая биномиальное распределение ошибок. Мы включили «участок» в качестве случайного эффекта, чтобы учесть возможность различий в пригодности участков для медоносных пчел. После этого предварительного этапа подгонки мы определили, что наша модель демонстрирует полное разделение (41). Поэтому мы подогнали последующие модели, используя коррекцию смещения (42,43) , которая в настоящее время применяется только для обобщенных линейных моделей (GLM). Мы включили местоположение как фиксированный эффект в наши окончательные модели, позволяющий отбросить этот член, если он будет признан неважным для присутствия/отсутствия колонии пчел во время выбора модели. Мы подобрали все подмножества полной модели и провели усреднение моделей для всех моделей ниже ΔAICc = 6 (40,44). Мы использовали R для анализа и построения графиков (39) с помощью нескольких пакетов (43,45,46,47,48,49,50,51,52,53,54,55).
Результаты
Одичавшие колонии A. mellifera были широко распространены в регионе исследования, и на всех участках, кроме двух (из 23), были обнаружены признаки использования колониями гнездовых ящиков (Дополнительная информация). Использование медоносными пчелами было зарегистрировано в 111 из 566 ящиков. В 26 случаях имелись свидетельства использования в прошлом, но живые колонии не присутствовали во время исследования, поэтому мы не включали эти записи в анализ. В 35 случаях гнездовые ящики были заняты в течение некоторого времени, но были покинуты до окончательного обследования. В остальных 50 случаях гнезда были заняты пчелами, и они все еще присутствовали в них во время окончательного обследования. Исследование с самым высоким уровнем использования гнездовых ящиков было зарегистрировано весной 2011 года, во время которого мы обнаружили 36 занятых гнездовых ящиков на площади 52,5 га, покрытой 10 крупными участками, выявив плотность 69 колоний на км2 леса. Доля гнездовых ящиков, занятых колониями медоносных пчел, была очень похожей в ходе исследования, проведенного год спустя, весной 2012 года.
В двух типах гнездовых ящиков, исключенных из уточненного набора данных, по крайней мере, в некоторых исследованиях находились пчелы. Первый (аналогичный типу B, но с отверстием диаметром 60 мм) был исключен из дальнейшего анализа, поскольку он был установлен только на одном участке (SAG) и имел мало записей (восемь). Пять из них были заняты по крайней мере в течение одного исследования, включая два случая, которые сохранялись в течение нескольких исследований. Другой тип ящиков (d = 170 мм, w = 170 мм, h = 500 мм, диаметр отверстия = 40 мм) был размещен 24 раза на двух остаточных участках, и живые пчелы были обнаружены в пяти различных ящиках в течение трех исследований, но ни один улей не сохранялся более одного периода исследований.
Свидетельства использования гнезд медоносными пчелами были зарегистрированы в 90 из 299 гнездовых ящиков из уточненного набора данных. Из них 45 ящиков (15,1%) были заняты пчелами и все еще присутствовали в последнем исследовании, а также 25 ящиков (8,3%), которые были заняты в течение некоторого времени, но затем были заброшены. В 20 случаях (6,6%) имелись признаки использования пчелами, но живые пчелы не присутствовали во время исследования, поэтому мы не включали их в статистический анализ. Заселение гнезд пчелами иногда приводило к тому, что гнездовые ящики пропадали из дальнейшего исследования: некоторые ящики разрушались под тяжестью улья, а другие пропадали при обстоятельствах, позволяющих предположить, что они были удалены людьми, стремящимися вернуть пчел к одомашниванию.
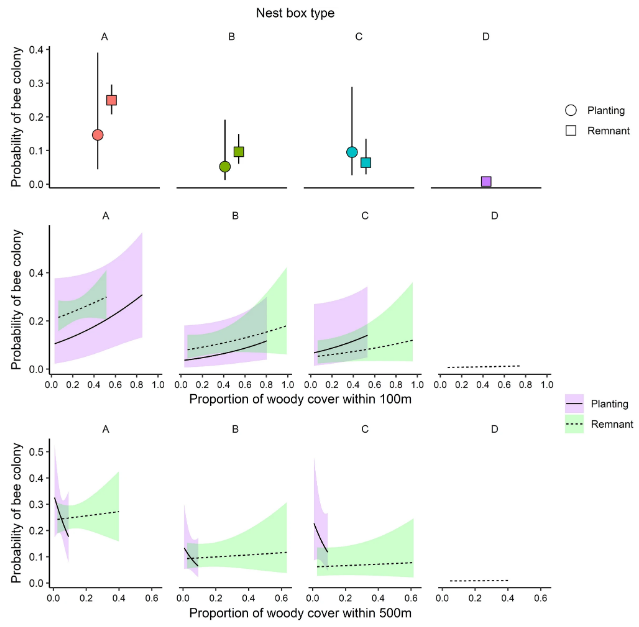
Если рассматривать исследование весной 2011 года, то гнездовой ящик типа А имел самую высокую вероятность быть занятым пчелами, и этот эффект был значительным при сравнении всех остаточных участков (p = 0,042; Таблица 4, Рис. 2). Доверительные интервалы были шире для оценок из посадок, где было меньше повторов. Было получено слабое доказательство интерактивного эффекта типа участка и доли древесного покрова в пределах 500 м (p = 0,091, Таблица 4), поскольку колонии чаще регистрировались в посадках, чем в остатках, когда древесный покров в пределах 500 м был очень низким, тогда как при увеличении древесного покрова картина менялась на противоположную. Важно отметить, что максимальный уровень древесного покрова на расстоянии 500 м был низким для насаждений и более высоким для остатков (рис. 2). Другим заметным членом в усредненной модели было взаимодействие древесного покрова в пределах 100 м и типа участка (т.е. посадки против остатков). Хотя вероятность использования следующего ящика, по-видимому, увеличивалась с ростом древесного покрова на расстоянии 100 м на обоих типах участков, ни одно из этих условий само по себе, ни эффект взаимодействия не были статистически значимыми.

Обсуждение
Широко распространенное и быстрое занятие искусственных гнездовых ящиков одичавшими колониями A. mellifera в этом регионе Австралии указывает на то, что до начала исследования существовала существующая и широко распространенная исходная популяция медоносных пчел, которая была достаточно энергичной, чтобы поддерживать рост популяции почти на всех обследованных нами участках. Возможные источники расширения популяции включают как ранее существовавшую одичавшую популяцию, так и управляемые ульи, которые в этом ландшафте часто размещаются рядом с полями канолы или рядом с цветущим Eucalyptus melliodora (56), распространенным местным деревом в регионе. Быстрое занятие гнездовых ящиков предполагает, что доступность нектара и пыльцы в ландшафте поддерживает рост популяции, но подходящие возможности для гнездования являются ограничивающим фактором.
Плотность колоний одичавших медоносных пчел, обнаруженных в данном исследовании (69 колоний на км2), является нижней границей истинной плотности, поскольку наши данные получены только из искусственных гнездовых ящиков. Тем не менее, даже этот уровень очень высок по сравнению с другими наземными исследованиями плотности колоний, в 5,5 раз выше, чем у Baum et al. (20), это самая высокая плотность, зарегистрированная за пределами Австралии, и сравнимая только с Oldroyd et al. (22). В совокупности наше исследование и исследование Ref. (22) позволяют предположить, что такая высокая плотность колоний не является необычной для Австралии. Учитывая, что наше исследование охватывает несколько лет повторных исследований (при одинаковой плотности в течение двух лет) и охватывает участки в широком регионе, наши результаты не могут быть отвергнуты как нерепрезентативные. Возможно, Австралия может поддерживать очень высокую плотность колоний одичавших медоносных пчел из-за обилия местных видов растений, богатых нектаром, и потому что паразитический клещ Varroa destructor еще не прижился в Австралии (57,58).
Исследования с использованием приманивания трутней и генетического анализа, включая некоторые из ландшафтов, близких к тому, где мы проводили данное исследование (59) , показали гораздо более низкую расчетную плотность колоний, чем мы. Однако эти исследования не следует напрямую сравнивать с нашим наземным исследованием. В дополнение к известным источникам заниженной оценки (рассмотренным в разделе «Введение»), в этих исследованиях по-разному определялась площадь выборки. Наше исследование включало только древесную растительность в качестве потенциального места гнездования, и мы рассчитывали площадь соответствующим образом. В отличие от этого, при исследованиях приманивания дронов выборки делаются из ландшафта, который включает потенциальные гнездовые местообитания, а также участки, где нет подходящих возможностей для гнездования медоносных пчел (например, поля, очищенные от всех деревьев). Тщательная интерпретация требует, чтобы используемая концепция плотности хорошо соответствовала интересующему вопросу. Если оценка Баума и др. (20) (12,5 колоний на км2) скорректирована с учетом того, что только 56% этого ландшафта составляла древесная растительность, способная обеспечить пчелам возможность гнездования, плотность колоний может быть пересчитана как 28 колоний на км2 древесной растительности. Даже при таком более высоком уровне плотность колоний, зарегистрированная в нашем исследовании, в 2,5 раза выше, чем в исследовании Баума и др. (20).
Достаточно ли высока плотность колоний, чтобы повлиять на местную экосистему?
Фрагменты леса, которые были в центре нашего исследования, находятся в более широком ландшафте, где древесная растительность была экстенсивно вырублена и заменена сельскохозяйственной системой с преобладанием земледелия и скотоводства (60). Хотя возможности гнездования ограничены фрагментами леса либо в естественных дуплах, либо в искусственных гнездовых ящиках, установленных на деревьях, пчелы могут добывать корм на многие километры (61) , поэтому высокая плотность в лесах поддерживается сбором пыльцы и нектара на территории, выходящей далеко за пределы небольших участков леса. Можно возразить, что воздействие пчел рассеяно (и, следовательно, разбавлено) на более обширной территории, чем лесные массивы, на которых мы сосредоточились, и что наша оценка плотности была бы ниже, если бы она была рассчитана в этом более крупном масштабе. Однако мы утверждаем, что поскольку фрагменты родных лесов являются очагами высокого биоразнообразия (62) , мы должны быть обеспокоены появлением такой высокой плотности колоний в этих участках, поскольку это, скорее всего, приведет к локальным последствиям, даже если популяция пчел использует ресурсы, собранные с более обширной территории.
Хотя многие исследования установили негативное воздействие одичавших медоносных пчел на местное биоразнообразие (6) , нет работ, связывающих масштаб этого воздействия с состоянием более широкой популяции медоносных пчел. Тот факт, что мы зафиксировали значения плотности колоний, превышающие все предыдущие исследования, за исключением одного, проведенного в южной части Австралии (22) , дает нам основание опасаться, что такая плотность популяции окажет значительное воздействие на местные виды, включая некоторые уязвимые. Однако единственный доступный подход, позволяющий сравнить масштабы воздействия на цветочные ресурсы и опыление, — это сравнение с рекомендуемыми нормами содержания ульев в сельском хозяйстве. При плотности 69 колоний на км2 местная плотность превышает некоторые рекомендации по норме содержания ульев для опыления малины, вики, различных дынь и бахчевых (17). В то время как местная плотность колоний медоносных пчел в наших фрагментированных лесах сравнима с этими сельскохозяйственными условиями, немногие местные растения будут производить цветы в масштабах, сравнимых с сельскохозяйственными культурами, а одичавшие колонии поддерживают круглогодичное присутствие, что совершенно не похоже на временное обеспечение управляемыми пчелами в пик сезона цветения в сельском хозяйстве. Поэтому можно ожидать, что обилие пчел по отношению к количеству цветов на наших участках будет превышать уровни в сельском хозяйстве большую часть времени. Можно ожидать, что такое высокое обилие пчел по отношению к количеству цветов повлияет на скорость опыления многих местных видов растений, которые привлекают A. mellifera (4) , и вызовет конкуренцию с местными животными, которые полагаются на те же ресурсы (7).
Воздействие обильных колоний медоносных пчел, эксплуатирующих цветы, является важным аспектом воздействия на биоразнообразие, однако конкуренция за гнезда оказывает влияние на другой и дополнительный набор видов. Уже установлено, что присутствие A. mellifera отталкивает другие виды от использования гнездовых ящиков в этом регионе, особенно виды позвоночных, для которых предназначены эти гнездовые ящики (25). Наш анализ подтверждает масштаб и степень этой проблемы. Поскольку мы сосредоточились на использовании искусственных гнездовых ящиков, наше исследование не дает прямой информации о конкуренции за естественные дупла, но быстрое освоение медоносными пчелами искусственных гнездовых ящиков соответствует пониманию того, что естественные дупла находятся в дефиците.
Учитывая широкое использование дупла деревьев пчелами A. mellifera, интересно изучить ценность рассеянных деревьев в сильно расчищенных ландшафтах для этого инвазивного вида. Наши данные показывают, что гнездовые ящики, размещенные в растительности, где вряд ли есть естественные дупла (т.е. в посадках), с наибольшей вероятностью будут заняты, когда в радиусе 500 м мало других деревьев (т.е. когда древесный покров низкий). Возможно, возможности для гнездования находятся на высоте в ландшафтах, где есть культуры, обеспечивающие ресурсы для медоносных пчел (например, рапс в нашем ландшафте 63), но при этом мало деревьев, которые могут поддерживать естественные дупла. Зрелые деревья на пастбищах, как известно, обеспечивают естественные дупла, которые в значительной степени востребованы другими видами фауны, зависящими от дупла, в этих ландшафтах (64) и предпочитаются некоторыми видами позвоночных (65).
Древесный покров и тип ящика влияют на заполняемость
Существует несколько способов, с помощью которых структура растительности (т.е. остатки леса или насаждения) и древесный покров могут влиять на выбор места для гнезда A. mellifera. Например, деревья могут предоставлять как естественные углубления для гнезд (что может уменьшить использование гнездовых ящиков), так и цветочные ресурсы для пчел (что может увеличить количество поддерживаемых колоний). Эти сложности особенно важны в нашем регионе исследования, где деревья рода Eucalyptus (а) широко распространены и доминируют в лесистых ландшафтах, (б) являются высоко ценимым кормом для медоносных пчел (56) и (в) известно, что в них много дупел (66). Поэтому неудивительно, что эти лесные и ландшафтные факторы были важными предикторами медоносных пчел в нашем анализе, и что их влияние было сложным и взаимодействующим.
Значительное влияние типа ящика на вероятность его заселения пчелами согласуется с результатами других исследований, которые указывают на важную роль конструкции гнездового ящика (31). Общий процент занятости гнездовых ящиков в нашем уточненном наборе данных составил 15,1%, но этот показатель включает один тип ящика, который не использовался (тип D), и другой, который был занят на 23,8% (тип A). Предпочтительная конструкция гнездового ящика (тип А) имела объем 14,5 л, что соответствует наблюдаемому диапазону естественных полостей, используемых медоносными пчелами, хотя и находится на низком уровне (67). Напротив, гнездовой ящик типа D имел объем 3,4 л, что меньше, чем наблюдается в естественных полостях, используемых пчелами (67) , что может объяснить, почему он не использовался одичавшими пчелами в нашем исследовании. Сили и Морс (67) также сообщают, что площадь гнездовых отверстий в естественных полостях, выбираемых медоносными пчелами, составляет от 1000 мм2 до 4000 мм2 , что указывает на то, что отверстия в ящиках типов B и C были больше, чем обычно предпочитаемые. Эти различия между гнездовыми ящиками в пригодности для A. mellifera необходимо учитывать при сравнении различных исследований. Инновации в конструкции гнездовых ящиков могут помочь уменьшить конфликт между различными пользователями. В то время как все типы ящиков в нашем исследовании были построены из дерева, недавнее исследование показало, что ящики из ПВХ меньше привлекают медоносных пчел и поэтому могут быть более доступны для позвоночной фауны (32).
Выводы
Обнаруженная нами высокая плотность одичавших колоний A. mellifera имеет значительные последствия для сохранения природы в этом ландшафте и указывает на аналогичный риск для других ландшафтов, где может развиться высокая плотность колоний. Воздействие на опыление будет сильным для многих видов растений, учитывая, что медоносные пчелы являются наиболее распространенными посетителями цветов в естественных местах обитания во всем мире, как в их родных, так и в инвазивных ареалах (4). Уже известно, что на сообщества местных пчел сильно влияют экологические фильтры, связанные с фрагментацией среды обитания (68) , поэтому присутствие большого количества инвазивных видов пчел добавит еще один сильный экологический фильтр. Многие животные, зависящие от дупла, которые могут конкурировать с колониями одичавших пчел за пространство, уже ограничены редкостью определенных типов дупла (69), а частое использование пчелами искусственных гнезд подрывает эффективность одного из наиболее распространенных природоохранных мероприятий, призванных смягчить эту проблему. Apis mellifera является исключительным видом в большинстве пчелиных сообществ из-за большого размера колоний. Это особенно верно в нашем регионе, где нет других эусоциальных видов пчел (70). Необходимы дальнейшие исследования, чтобы определить, может ли высокая плотность колоний одичавших медоносных пчел, зафиксированная в данном исследовании, встречаться и во многих других регионах мира, о которых известно, что там есть популяции одичавших медоносных пчел, но по которым до сих пор нет данных исследований.
Ссылки:
1.Geslin, B. et al. Massively introduced managed species and their consequences for plant–pollinator interactions. Adv. Ecol. Res. 57, 147–199 (2017). Article Google Scholar
2.Huryn, V. M. B. Ecological impacts of introduced honey bees. Q. R. Biol. 72, 275–297 (1997). Article Google Scholar
3.Stout, J. C. & Morales, C. L. Ecological impacts of invasive alien species on bees. Apidologie 40, 388–409 (2009). Article Google Scholar
4.Hung, K.-L.J., Kingston, J. M., Albrecht, M., Holway, D. A. & Kohn, J. R. The worldwide importance of honey bees as pollinators in natural habitats. Proc. R. Soc. Ser. B 285, 20172140 (2018). PDF opens in a new tab Article Google Scholar
5.Paini, D. R. Impact of the introduced honey bee (Apis mellifera) (Hymenoptera: Apidae) on native bees: A review. Austral Ecol. 29, 399–407 (2004). Article Google Scholar
6.Moritz, R. F. A., Hartel, S. & Neumann, P. Global invasions of the western honey bee (Apis mellifera) and the consequences for biodiversity. Ecoscience 12, 289–301 (2005). Article Google Scholar
7.Paini, D. R. & Roberts, J. D. Commercial honey bees (Apis mellifera) reduce the fecundity of an Australian native bee (Hylaeus alcyoneus). Biol. Cons. 123, 103–112 (2005). Article Google Scholar
8.Munoz, I. & De la Rua, P. Wide genetic diversity in old world honey bees threatened by introgression. Apidologie 52, 200–217 (2021). PDF opens in a new tab Article Google Scholar
9.Williams, I. H. The dependences of crop production within the European Union on pollination by honey bees. Agric. Zool. Rev. 6, 229–257 (1994). Google Scholar
10.Thompson, C. E., Biesmeijer, J. C., Allnutt, T. R., Pietravalle, S. & Budge, G. E. Parasite pressures on feral honey bees (Apis mellifera sp.). PLoS One 9, e105164 (2014). Article ADS PubMed Pub Med Central Google Scholar
11.Belsky, J. & Joshi, N. K. Impact of biotic and abiotic stressors on managed and feral bees. Insects 10, 233 (2019).
Article PubMed Central Google Scholar
12.Medina-Flores, C. A., Guzman-Novoa, E., Hamiduzzaman, M. M., Arechiga-Flores, C. F. & Lopez-Carlos, M. A. Africanized honey bees (Apis mellifera) have low infestation levels of the mite Varroa destructor in different ecological regions in Mexico. Genet. Mol. Res. 13, 7282–7293 (2014). Article CAS PubMed Google Scholar
13.Portman, Z. M., Tepedino, V. J., Tripodi, A. D., Szalanski, A. L. & Durham, S. L. Local extinction of a rare plant pollinator in Southern Utah (USA) associated with invasion by Africanized honey bees. Biol. Invasions 20, 593–606 (2018).
PDF opens in a new tab Article Google Scholar
14.Santos, G. M. D. et al. Invasive Africanized honeybees change the structure of native pollination networks in Brazil. Biol. Invasions 14, 2369–2378 (2012). PDF opens in a new tab Article Google Scholar
15.Chapman, R. E. & Bourke, A. F. G. The influence of sociality on the conservation biology of social insects. Ecol. Lett. 4, 650–662 (2001). Article Google Scholar
16.Aizen, M. A. et al. When mutualism goes bad: Density-dependent impacts of introduced bees on plant reproduction. New Phytol. 204, 322–324 (2014). Article Google Scholar
17.Breeze, T. D. et al. Agricultural policies exacerbate honeybee pollination service supply-demand mismatches across Europe. PLoS One 9, e82996 (2014). Article ADS PubMed PubMed Central Google Scholar
18.Baum, K. A. et al. Spatial distribution of Africanized honey bees in an urban landscape. Landsc. Urban Plan. 100, 153–163 (2011). Article Google Scholar
19.Ratnieks, F. L. W., Piery, M. A. & Cuadriello, I. The natural nest and nest density of the africanized honey-bee (Hymenoptera, Apidae) near Tapachula, Chiapas, Mexico. Can. Entomol. 123, 353–359 (1991). Article Google Scholar
20.Baum, K. A., Rubink, W. L., Pinto, M. A. & Coulson, R. N. Spatial and temporal distribution and nest site characteristics of feral honey bee (Hymenoptera: Apidae) colonies in a coastal prairie landscape. Environ. Entomol. 33, 727–739 (2004). Article Google Scholar
21.Rangel, J. et al. Africanization of a feral honey bee (Apis mellifera) population in South Texas: Does a decade make a difference?. Ecol. Evol. 6, 2158–2169 (2016). PDF opens in a new tab Article PubMed PubMed Central Google Scholar
22.Oldroyd, B. P., Thexton, E. G., Lawler, S. H. & Crozier, R. H. Population demography of Australian feral bees (Apis mellifera). Oecologia 111, 381–387 (1997). PDF opens in a new tab Article ADS CAS PubMed Google Scholar
23.Arundel, J. et al. Remarkable uniformity in the densities of feral honey bee Apis mellifera Linnaeus, 1758 (Hymenoptera: Apidae) colonies in South Eastern Australia. Austral Entomol. 53, 328–336 (2014). Article Google Scholar
24.Remm, J. & Lõhmus, A. Tree cavities in forests—The broad distribution pattern of a keystone structure for biodiversity. For. Ecol. Manag. 262, 579–585 (2006). Article Google Scholar
25.Lindenmayer, D., Crane, M., Blanchard, W., Okada, S. & Montague-Drake, R. Do nest boxes in restored woodlands promote the conservation of hollow-dependent fauna?. Restor. Ecol. 24, 244–251 (2016). Article Google Scholar
26.New South Wales Department of Planning, Industry and Environment 2003.
https://www.environment.nsw.gov.au/topics/animals-and-plants/threatened-species/nsw-threatened-species-scientific-committee/determinations/final-determinations/2000-2003/competition-from-feral-honeybees-key-threatening-process-listing (accessed 22 Feb 2021).
27.Goldingay, R. L., Rohweder, D. & Taylor, B. D. Nest box contentions: Are nest boxes used by the species they target?. Ecol. Manag. Restor. 21, 115–122 (2020). PDF opens in a new tab Article Google Scholar
28.Lindenmayer, D. B. et al. Are nest boxes a viable alternative source of cavities for hollow-dependent animals? Long-term monitoring of nest box occupancy, pest use and attrition. Biol. Cons. 142, 33–42 (2009). Article Google Scholar
29.Lindenmayer, D. B. et al. The anatomy of a failed offset. Biol. Conserv. 210, 286–292 (2017). Article Google Scholar
30.Macak, P. V. Nest boxes for wildlife in Victoria: An overview of nest box distribution and use. Vic. Nat. 137, 4–14 (2020). Google Scholar
31.Le Roux, D. S. et al. Effects of entrance size, tree size and landscape context on nest box occupancy: Considerations for management and biodiversity offsets. For. Ecol. Manag. 366, 135–142 (2016). Article Google Scholar
32.Berris, K. K. & Barth, M. PVC nest boxes are less at risk of occupancy by feral honey bees than timber nest boxes and natural hollows. Ecol. Manag. Restor. 21, 155–157 (2020). PDF opens in a new tab Article Google Scholar
33.Jaffe, R. et al. Estimating the density of honeybee colonies across their natural range to fill the gap in pollinator decline censuses. Conserv. Biol. 24, 583–593 (2010). Article PubMed Google Scholar
34.Utaipanon, P., Schaerf, T. M. & Oldroyd, B. P. Assessing the density of honey bee colonies at ecosystem scales. Ecol. Entomol. 44, 291–304 (2019). PDF opens in a new tab Article Google Scholar
35.Utaipanon, P., Holmes, M. J., Chapman, N. C. & Oldroyd, B. P. Estimating the density of honey bee (Apis mellifera) colonies using trapped drones: Area sampled and drone mating flight distance. Apidologie 50, 578–592 (2019).
PDF opens in a new tab Article CAS Google Scholar
36.Williamson, E. M. Reliability of honey bee hive density estimates using drone sampling: does relative hive size or distance affect a colony’s drone contribution? Honours Thesis, The University of Adelaide (2020).
37.Benson, J. S. The effect of 200 years of European settlement on the vegetation and flora of New South Wales. Cunninghamia 2, 343–370 (1991). Google Scholar
38.New South Wales Office of Environment and Heritage 2015. Upgraded NSW woody vegetation extent for 2011. http://data.auscover.org.au/xwiki/bin/view/Product+pages/nsw+5m+woody+extent+and+fpc (accessed 13 May 2020).
39.R Core Team. R: A Language and Environment for Statistical Computing. (R Foundation for Statistical Computing, 2020).
www.R-project.org (accessed 12 January 2021).
40.Burnham, K. P. & Anderson, D. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach (Springer, 2002). MATH Google Scholar
41.Albert, A. & Anderson, J. A. On the existence of maximum likelihood estimates in logistic regression models. Biometrika 71, 1–10 (1984). PDF opens in a new tab Article MathSciNet MATH Google Scholar
42.Firth, D. Bias reduction of maximum likelihood estimates. Biometrika 80, 27–38 (1993).
PDF opens in a new tab Article MathSciNet MATH Google Scholar
43.Kosmidis, I., Pagui, E. C. K. & Sartori, N. Mean and median bias reduction in generalized linear models. Stat. Comput. 30, 43–59 (2020). PDF opens in a new tab Article MathSciNet MATH Google Scholar
44.Anderson, D. R. Model Based Inference in the Life Sciences: A Primer on Evidence (Springer Science & Business Media, 2007).
Google Scholar
45.Barton, K. MuMIn: Multi-model inference. R package version 1.43.17 (2016).
46.Hijmans, R. J. Raster: Geographic Data Analysis and Modeling. R package version 3.4-5 (2020).
47.Kassambara, A. ggpubr: ‘ggplot2’ Based Publication Ready Plots. R package version 0.3.0 (2018).
48.Kosmidis, I. brglm2: Bias Reduction in Generalized Linear Models. R package version 0.6.2 (2020).
49.Kosmidis, I., Schumacher, D. detectseparation: Detect and Check for Separation and Infinite Maximum Likelihood Estimates. R package version 0.1 (2020).
50.Pebesma, E. Simple features for R: Standardized support for spatial vector data. R J. 10, 439–446 (2018). Article Google Scholar
51.Pateiro-Lopez, B., Rodriguez-Casal, A. Alphahull: Generalization of the Convex Hull of a Sample of Points in the Plane. R package version 2.2 (2019).
52.Wickham, H. ggplot2: Elegant Graphics for Data Analysis (Springer, 2009). Book MATH Google Scholar
53.Wickham, H. The split-apply-combine strategy for data analysis. J. Stat. Softw. 40, 1–29 (2011). Article Google Scholar
54.Wickham, H. Forcats: Tools for working with categorical variables (factors). R package version 0.5.0 (2018).
55.Wickham, H., François, R., Henry, L., Müller, K. dplyr: A Grammar of Data Manipulation. R package version 1.0.0 (2021).
56.Birtchnell, M. J. & Gibson, M. Long-term flowering patterns of melliferous Eucalyptus (Myrtaceae) species. Aust. J. Bot. 54, 745–754 (2006). Article Google Scholar
57.Steinhauer, N. et al. Drivers of colony losses. Curr. Opin. Insect Sci. 26, 142–148 (2018). Article PubMed Google Scholar
58.Cunningham, S. A., Heard, T. & FitzGibbon, F. The future of pollinators for Australian Agriculture. Aust. J. Agric. Res. 53, 893–900 (2002). Article Google Scholar
59.Hinson, E. M., Duncan, M., Lim, J., Arundel, J. & Oldroyd, B. P. The density of feral honey bee (Apis mellifera) colonies in South East Australia is greater in undisturbed than in disturbed habitats. Apidologie 46, 403–413 (2015).
PDF opens in a new tab Article Google Scholar
60.McIntyre, S. Ecological and anthropomorphic factors permitting low-risk assisted colonization in temperate grassy woodlands. Biol. Conserv. 144, 1781–1789 (2011). Article Google Scholar
61.Steffan-Dewenter, I. & Kuhn, A. Honeybee foraging in differentially structured landscapes. Proc. R. Soc. B Biol. Sci. 270, 569–575 (2003). Article Google Scholar
62.Wintle, B. A. et al. Global synthesis of conservation studies reveals the importance of small habitat patches for biodiversity. Proc. Natl. Acad. Sci. U.S.A. 116, 909–914 (2019). Article CAS PubMed Google Scholar
63.Arthur, A. D., Li, J., Henry, S. & Cunningham, S. A. Influence of woody vegetation on pollinator densities in oilseed Brassica fields in an Australian temperate landscape. Basic Appl. Ecol. 11, 406–414 (2010). Article Google Scholar
64.Lindenmayer, D. B. et al. New policies for old trees: Averting a global crisis in a keystone ecological structure. Conserv. Lett. 7, 61–69 (2014). Article Google Scholar
65.Crane, M. J., Lindenmayer, D. B. & Cunningham, R. B. The value of countryside elements in the conservation of a threatened arboreal marsupial Petaurus norfolcensis in agricultural landscapes of south-eastern Australia—the disproportional value of scattered trees. PLoS One 9, e107178 (2014). Article ADS PubMed PubMed Central Google Scholar
66.Gibbons, P., Lindenmayer, D. B., Barry, S. C. & Tanton, M. T. Hollow selection by vertebrate fauna in forests of southeastern Australia and implications for forest management. Biol. Conserv. 103, 1–12 (2002). Article Google Scholar
67.Seeley, T. D. & Morse, R. A. The nest of the honey bee (Apis mellifera L.). Insectes Soc. 23, 495–512 (1976). Article Google Scholar
68.Hung, K. L. J., Ascher, J. S., Davids, J. A. & Holway, D. A. Ecological filtering in scrub fragments restructures the taxonomic and functional composition of native bee assemblages. Ecology 100, e02654 (2019).
PDF opens in a new tab Article PubMed Google Scholar
69.Cockle, K. L., Martin, K. & Drever, M. C. Supply of tree-holes limits nest density of cavity-nesting birds in primary and logged subtropical Atlantic forest. Biol. Conserv. 143, 2851–2857 (2010). Article Google Scholar
70.Heard, T. Stingless bees. In Australian Native Bees: A Practical Hand Book 106–139 (NSW Department of Primary Industries, 2016).
71.Geoscience Australia 2006. GEODATA TOPO 250K. Commonwealth of Australia. http://pid.geoscience.gov.au/dataset/ga/63999 (accessed 11 December 2020).
Добавить комментарий