
Аннотация
Управляемые колонии медоносных пчел (Apis mellifera) содержатся с гораздо большей плотностью, чем естественные одичавшие или дикие колонии, что может иметь пагубные последствия для здоровья и выживания колоний, распространения болезней и поведения дрейфа (перемещения пчел между родными и неродными колониями). Мы оценили влияние простого вмешательства в управление пасекой (изменение плотности и внешнего вида колоний) на здоровье колоний. В частности, мы создали три конфигурации пасек «высокая плотность / высокий дрейф» («HD») и три конфигурации «низкая плотность / низкий дрейф» («LD»), каждая из которых состояла из восьми колоний. Ульи в конфигурации пасеки HD были одного цвета и размещены на расстоянии 1 м друг от друга в единый линейный массив, в то время как ульи в конфигурации пасеки LD были размещены на расстоянии 10 м друг от друга на разной высоте, обращены наружу по кругу и визуально различались цветами и символами, чтобы уменьшить случайный дрейф между колониями. Мы исследовали передачу заболеваний и динамику между конфигурациями пасек путем очистки всех колоний от паразитического клеща Varroa destructor и последующей инокуляции двух случайно выбранных колоний на каждой пасеке контролируемыми дозами клеща. Мы наблюдали за колониями в течение двух лет и обнаружили, что на пасеке с конфигурацией LD значительно увеличилось производство меда и снизилась смертность в зимний период. Инокуляция и вмешательство в управление пасекой взаимодействовали, влияя на уровень расплодного клеща, причем самый высокий уровень был в инокулированных колониях в конфигурации HD. Наконец, вероятность дрейфа фуражиров в конфигурации пасеки HD была более чем в три раза выше. Наши результаты показывают, что относительно простое изменение в управлении — размещение колоний в визуально сложных кругах с низкой плотностью, а не в линейных массивах с высокой плотностью — может обеспечить значительные преимущества для здоровья и продуктивности управляемых колоний медоносных пчел.
Введение
Широкую озабоченность вызывает продолжающееся сокращение численности управляемых медоносных пчел (Apis mellifera). Например, по оценкам, с 1941 по 2008 год количество управляемых колоний в США сократилось на 61% из-за социально-экономических факторов, вызывающих высокую текучесть колоний и повышенную смертность [1,2]. Управляемые колонии обычно содержатся в непосредственной близости и их плотность на много порядков выше, чем у их одичавших или диких собратьев, плотность которых обычно колеблется в пределах 1-6 колоний на км2 [3,4]. Управляемые пасеки, напротив, расположены плотно, колонии обычно находятся на расстоянии ≤1 м друг от друга [5] и могут содержать 40 колоний на небольшом пространстве. Такое резкое изменение плотности может иметь серьезные последствия для здоровья и выживания колоний, передачи болезней и поведения при дрейфе (т.е. когда пчелы заходят в нерожденную колонию).
Плотность популяции изучалась как ключевой фактор экологических отношений еще Мальтусом [6], который впервые описал зависимость смертности и плодовитости от плотности. Плотность известна как важный фактор динамики популяций во многих таксонах, включая насекомых [7], рыб [8], растения [9] и млекопитающих [10]. Плотность также может быть важным модулятором других экологических факторов, включая ландшафтные модели, такие как размер участка [11] и экологические взаимодействия, такие как хорошо изученное влияние плотности добычи на уровень потребления хищниками [12]. Исследования других видов социальных насекомых показали, что конкуренция за кормовое пространство отражается на том, насколько близко распределены муравьиные колонии [13], а рабочие муравьи в переполненных колониях тратят больше энергии, что может повлиять на производительность и приспособленность колонии [14].
В сельскохозяйственных системах распространены высокие плотности скота, и скученность может иметь негативные последствия для производительности животных. Например, высокая плотность поголовья может увеличить регуляцию генов стресса и снизить регуляцию иммунных генов у рыб [15], в то время как коровы в условиях высокой плотности содержания уменьшают количество времени, которое они тратят на кормление [16]. У медоносных пчел скученность внутри колонии может оказывать пагубное влияние на продуктивность колонии и продолжительность жизни пчел [17,18]. Скученность в кормовых условиях может также инициировать сигналы о прекращении кормовой деятельности или уменьшить привлечение новых фуражиров, тем самым снижая эффективность кормовой деятельности колонии [19,20]. Кроме того, межколониальная скученность может оказывать пагубное влияние на ошибки при наведении пчел-трутней и увеличивать нагрузку паразитов [5]. Напротив, пасеки с низкой плотностью могут оказывать негативное влияние на частоту спаривания для полиандровых маток медоносных пчел [21].
Как теоретические, так и эмпирические экологические исследования показывают, что плотность популяции также является ключевым фактором, определяющим экологию и динамику заболеваний [22,23]. Экология болезней предсказывает, что более высокая плотность популяций хозяев и большее смешение популяций хозяев приведет к большей передаче болезней и увеличению бремени болезней [24] и может привести к эволюции повышенной вирулентности паразитов [25]. В системе медоносной пчелы предполагается, что увеличение плотности колоний и передача инфекции, вероятно, способствуют эволюции вирулентности патогенов и вредителей [26]. Паразитарный компонент данного исследования сосредоточен на облигатном эктопаразитическом клеще Varroa destructor, поскольку он считается самой большой биотической угрозой для медоносных пчел [27]. Было также показано, что паразитизм V. destructor влияет на способность медоносных пчел к хомингу и может влиять на поведение дрейфующих пчел и распространение болезней [28].
При типичной планировке пасеки колонии размещаются близко друг к другу, выстраиваются в ряд с входами, обращенными в одном направлении, и окрашиваются в один цвет [5]. При такой системе управления наблюдается высокая плотность колоний медоносных пчел, скученность кормилиц и значительный уровень смешивания пчел между этими колониями. Такое смешение может происходить намеренно (перенос рамок для выравнивания силы колонии) или непреднамеренно, за счет более высоких показателей дрейфа пчел в визуально похожие колонии [5,29]. Учитывая, что все эти факторы согласуются с негативным влиянием плотности и конфигурации пасеки [5], мы предположили, что стандартные методы управления пчеловодством усилят конкуренцию за цветочные ресурсы между колониями, приведут к увеличению бремени болезней и их передачи, а также негативно повлияют на здоровье, продуктивность и выживание колоний. В частности, мы предсказывали, что уровень V. destructor на пасеках с высокой плотностью будет расти быстрее и поддерживать более высокое бремя клещей на протяжении всего эксперимента по сравнению с пасеками с низкой плотностью. Кроме того, мы ожидали, что колонии в конфигурациях с высокой плотностью будут иметь более низкую силу колонии (измеряемую по производству взрослых особей и расплода, а также по производству меда) и выживаемость, а также более высокую нагрузку V. destructor, передачу V. destructor и дрейф рабочих пчел. Наконец, мы ожидали наблюдать более высокие показатели дрейфа на пасеках с конфигурацией высокой плотности по сравнению с пасеками с конфигурацией низкой плотности.
Материалы и методы
Обзор
Чтобы определить влияние конфигурации пасеки медоносных пчел (сочетание расстояния между колониями, их расположения и визуальной сложности) на здоровье колоний, бремя паразитов и дрейф пчел, мы провели двухлетнее исследование, в котором сравнивали линейную конфигурацию колоний с высокой плотностью (высокая плотность, далее «HD») с круговой конфигурацией с низкой плотностью (низкая плотность, далее «LD»). Мы создали три конфигурации пасек HD и три конфигурации пасек LD в июне 2015 года в окрестностях Афин, штат Джорджия, США, обслуживаемых Лабораторией медоносных пчел Университета Джорджии. Каждая пасека находилась на расстоянии не менее 3 км от ближайшей пасеки и состояла из восьми колоний, изначально размещенных в стандартных пятирамочных нуклеусах Лангстрота, в общей сложности 48 колоний. Мы расположили колонии HD в линейный массив с расстоянием 1 м между колониями и со всеми входами, обращенными в одном направлении (рис. 1A). Все колонии HD были окрашены в белый цвет (т.е. не было цветовой вариативности) и размещены на одинаковой высоте (200 см) над землей. Мы выбрали круговое расположение колоний на пасеке LD с расстоянием 10 м между колониями и всеми входами, обращенными наружу от центра круга (Рис. 1B). Чтобы максимизировать способность пчел визуально различать колонии, мы покрасили колонии LD в разные цвета, нарисовали разные символы на их входах и разместили колонии на трех разных высотах над землей (6, 200, 400 см, причем пространственно ближайшие колонии находились на разных высотах). Сначала мы очистили все колонии от V. destructor, а затем инокулировали две случайно выбранные колонии на пасеке 200 взрослыми клещами, чтобы отразить естественное заражение клещами. Мы наблюдали за колониями в течение двух лет.

A показана конфигурация HD, а B — конфигурация LD. Каждая конфигурация была повторена 3 раза. Направление стрелки указывает на вход в колонию.
Затем мы оценили влияние различных конфигураций пасеки (HD против LD), экспериментально скрещенных с инфекцией V. destructor, на четыре аспекта здоровья медоносных пчел: 1) сила колонии (измеряемая численностью взрослых пчел, расплода и производством меда); 2) выживаемость колонии; 3) размножение и распространение V. destructor; и 4) поведение рабочих пчел в дрейфе.
Сила колонии и зараженность клещами
Чтобы свести к минимуму вариации, мы начали с высоко стандартизированных колоний. В каждую колонию мы поместили по спаренной матке и 1,1 кг (2,5 фунта) взрослых пчел, вытряхнутых в пакет. Все королевы были от одного и того же заводчика в Южной Джорджии, США, а взрослые пчелы — из общих источников. Пакеты обрабатывали от V. destructor двумя разными методами, чтобы обеспечить максимальную очистку от клещей. Сначала мы использовали метод сахарной пудры [30], чтобы стимулировать уход за пчелами и вытеснение клещей. Затем пакеты были помещены в темное помещение на ночь при температуре 16,6°C (62°F) и опрысканы сахарной водой за час до нанесения 30 мл раствора щавелевой кислоты [31]. Каждый пакет был установлен через три дня на случайно выбранной пасеке. Первоначальная липкая доска была оставлена в каждой колонии на 72 часа, чтобы убедиться, что колонии свободны от клещей Varroa destructor. Чтобы изучить совместное влияние плотности пасеки, расположения колоний и визуальной сложности пасеки на динамику паразитов, мы случайным образом выбрали две колонии на каждой пасеке и инокулировали эти колонии 200 взрослыми клещами V. destructor. Клещей собирали из исходных колоний вне эксперимента, просеивая сахарную пудру над колонией и собирая смещенных клещей на дне колонии. Мы использовали маленькие кисточки из натурального волокна, чтобы поместить клещей на влажные кофейные фильтры. Мы держали клещей в инкубаторе при температуре 35°C (95°F), пока не были собраны все клещи. Затем мы равномерно переносили всех клещей на открытую расплодную рамку и ждали одну минуту, чтобы убедиться, что клещи прикрепились к восковым клеткам колонии. Мы подчеркиваем, что инокулированные колонии ближе к обычным условиям пчеловодства (т.е. бремя клещей V. destructor, которое можно встретить на обычной пасеке), чем обработка с почти полной очисткой в нашем эксперименте, поскольку динамика популяции V. destructor находится в состоянии «постоянного обмена» в районах с высокой плотностью колоний [27]. В большом шестилетнем исследовании Трейнор и др. [32] обнаружили, что нередко в колониях пчеловодов весной уровень клещей не равен нулю. В таких условиях практически невозможно поддерживать нулевой уровень клещей в колониях, как показали Делаплан и Худ [33], которые сообщили о средней численности клещей в августе 111±69 в колониях, в которых некоторые колонии на пасеке постоянно обрабатывались от клещей, а другие нет. Для того, чтобы не отвлекаться на эти первоначальные колонии (и их маток), мы ввели контроль роя на колониях, которые могли роиться, разделив эти колонии. Мы стандартизировали контроль роя, чтобы не подвергать риску маленькие колонии. В общей сложности 38 из 48 колоний были разделены для предотвращения роения, и все это произошло в марте 2016 года. Мы использовали точный тест Фишера, чтобы определить, что не было значительной разницы (P = 0,29) в количестве разделений между нашими группами. Во время эксперимента мы не проводили никаких мер борьбы с V. destructor. Колонии были переселены в 10-рамочные ульи и подселены в соответствии с потребностями роста и места для хранения. Мы кормили колонии раствором сиропа — обычная практика в пчеловодстве, которую мы стандартизировали, давая одинаковые объемы всем колониям независимо от потребности. Мы продолжали эксперимент с июня 2015 года по май 2017 года, когда из первоначальных 48 колоний в живых осталось только 12.
Сбор данных
Оценка силы колонии.
Чтобы определить влияние плотности и расположения пасеки на здоровье колонии, мы проводили периодические измерения здоровья на протяжении всего эксперимента. Мы следовали рекомендациям по оценке силы колонии, описанным в работе Делаплана и других [34], чтобы измерить численность взрослых пчел, количество расплода и запас меда для каждой колонии. Мы проводили эту оценку колоний семь раз в течение двух лет эксперимента. Мы также записали дату, когда каждая колония была признана мертвой (без живых пчел), и последнюю известную дату, когда она была жива, для анализа выживаемости.
Измерение зараженности V. destructor.
Мы измеряли уровень заражения V. destructor тремя различными способами. Во-первых, мы использовали метод промывки спиртом, описанный Фризом и другими [35]. Этот метод включает уничтожение 300 пчел из колонии в спирте и подсчет пчел и клещей (которые отделяются от пчел, что облегчает подсчет) для получения относительного уровня клещей в популяции взрослых пчел. Промывки спиртом проводились примерно ежемесячно летом и осенью и каждые 3 месяца зимой и весной. В течение всего эксперимента было взято девять проб спиртовых смывов. Во-вторых, мы использовали липкие доски, оставленные в колонии на 72 часа [36], стандартный метод оценки уровня V. destructor в колонии путем сбора клещей, которые падают и попадают в ловушку на доске, размещенной на дне колонии. Мы измеряли уровень клещей с помощью липких досок семь раз в течение первого года эксперимента, включая один раз сразу после установки пакета, чтобы убедиться, что колонии свободны от V. destructor. В течение первого года липкие доски снимали примерно раз в два месяца, а во второй год эксперимента по логистическим причинам не продолжали. В-третьих, мы измерили популяцию клещей в выводковых ячейках, открыв 100 закрытых выводковых ячеек в каждой колонии и подсчитав количество клещей [37]. Клещей в выводковых ячейках исследовали примерно с интервалом в 4 месяца. Мы измеряли уровень клещей в выводковых ячейках шесть раз в течение эксперимента.
Дрейфующее поведение
.Для количественной оценки потенциального влияния планировки пасеки на поведение пчел в дрейфе мы пометили отдельных пчел уникальными пронумерованными метками и использовали видеокамеры, расположенные над входом в колонию, чтобы зафиксировать вхождение и выход пчел [38].Мы провели мечение и видеосъемку в сентябре 2015 года. Мы помечали только что появившихся рабочих, чтобы убедиться, что помеченные пчелы происходят из этой колонии. Мечение было разделено на три последовательные недели. Каждую неделю мы помечали до 100 пчел во всех колониях с одной пасеки HD и одной пасеки LD. В дни 24 и 25 после мечения мы записывали от пяти до семи часов видео в часовых сегментах в каждой колонии. Вместе с коллегами по компьютерным наукам мы разработали программу анализа видео в Matlab под названием GRAPHITE для изучения каждого кадра видео и извлечения кадров, содержащих метки [38]. С помощью этой программы мы идентифицировали метки на видео и определили: из какой колонии пчела с меткой; входила ли пчела или выходила; и несла ли пчела пыльцу.
Статистический анализ
Обзор
Мы исследовали влияние двух основных объясняющих переменных на несколько показателей здоровья медоносных пчел, описанных выше в разделе сбора данных. Объясняющими переменными были: (1) конфигурация пасеки (комплексная мера плотности пасеки, расположение колоний, цвет и символ колонии); (2) обработка от паразитов (очищенная против привитой: была ли колония привита стандартной инфекцией V. destructor в начале эксперимента). Мы провели три общих класса анализов, основанных на следующих переменных ответа: 1) зараженность клещами на уровне колонии и параметры здоровья колонии; 2) выживаемость на уровне колонии; и 3) дрейфующее поведение.
Сила колонии и зараженность клещами.
Продольные повторные измерения и вложенные конструкции, использованные в нашем эксперименте, могут привести к временной и внутрисубъектной автокорреляции, что нарушает предположения о независимости для параметрических и линейных методов регрессии. Поэтому мы использовали обобщенные уравнения оценки (GEE) для учета повторяющихся измерений, включая автокорреляцию. Мы использовали функцию ‘geeglm’ из пакета ‘geepack’ v.1.2-1 [39] в R v.3.4.2 [40] для определения и оценки моделей GEE, поскольку она позволяет использовать продольные данные с отсутствующими наблюдениями. Мы блокировали данные по пасекам и колониям и использовали авторегрессионную (AR1) автокорреляционную структуру. Каждая исходная модель была определена с использованием двух объясняющих переменных и их взаимодействия. В тех случаях, когда GEE не сходился, к каждой дате выборки применялся тест Wilcoxon signed-rank и процедура Benjamini-Hochberg для корректировки коэффициента ложного обнаружения при тестировании множественных сравнений.
Анализ выживаемости
Мы провели анализ выживаемости колоний с использованием обеих объясняющих переменных и их взаимодействия, конфигурации пасеки и статуса инокуляции колоний. Мы также провели отдельный анализ выживаемости в зимний период, поскольку инфекция V. destructor связана со снижением выживаемости в зимний период [41,42]. Колонии периодически осматривались на протяжении всего эксперимента, и точное время гибели колоний определить было невозможно. Поэтому мы использовали интервал между датой наблюдаемой гибели колонии и датой последней известной жизнеспособности колонии. Учитывая такую структуру данных, мы проанализировали выживаемость с помощью модели пропорциональной опасности Кокса с интервальным цензурированием с помощью пакета «frailtypack» [43] в R, который позволяет задать эквивалент случайных эффектов (только для пасеки, поскольку выживаемость одной колонии во времени является переменной ответа). Поскольку данные по выживаемости в зимний период состояли исключительно из биномиальных данных в одной временной точке (т.е. выжили и не выжили зимой), мы использовали отдельную биномиально-ошибочную обобщенную линейную смешанную модель (GLMM) с помощью пакета lme4 [44] в R со случайными эффектами для идентичности колоний, вложенными в пасеку. Мы проверили модель на чрезмерную дисперсию.
Анализ дрейфующих данных
Чтобы понять, как статус инокулированной колонии и конфигурация пасеки влияют на поведение пчел при дрейфе, мы провели GLMM с использованием пакета lme4, при этом количество событий дрейфа моделировалось ошибками Пуассона. Модель была проверена на чрезмерную дисперсию. Дрейф может также коррелировать с близко расположенными соседями. Мы использовали коррелограммы Мантеля для оценки корреляции между дрейфом и относительным положением колонии (ближайший сосед, второй ближайший сосед, третий ближайший сосед и т.д.) с помощью пакета ‘vegan’ в R [45], используя прогрессивную поправку для множественных сравнений.
Результаты
Обзор
На протяжении всего эксперимента мы собирали обширные данные о силе колоний, уровне клещей и перемещении отдельных пчел. В результате оценки состояния колонии было проведено 202 измерения: популяции взрослых пчел, покрытия расплода и хранения меда. Для оценки уровня V. destructor на протяжении всего эксперимента мы собрали 316 липких досок, 279 спиртовых смывов (каждый из которых содержал примерно 300 рабочих пчел) и 208 подсчетов клещей в расплоде (каждый из которых включал 100 расплодных ячеек). Мы записали и использовали программу GRAPHITE для обработки около 1200 часов видео, отслеживая индивидуально помеченных пчел. Мы наблюдали 120 уникальных меченых пчел в 242 отдельных момента времени.
Сила колонии
GEE-модель хранения меда не сходилась, вероятно, из-за очень сильной сезонной закономерности в производстве меда. После применения процедуры множественных сравнений Бенджамина-Хохберга для каждой из дат отбора проб был проведен знаково-ранговый тест Уилкоксона, и оказалось, что в конфигурациях LD было значительно больше запасов меда в четыре из пяти дат отбора проб (P = 0,020, 0,010, 0,040; рис. 2 и S1A рис.), а единственной не значимой датой отбора была первая проба. Ни одна из моделей GEE для популяции взрослых пчел и количества расплода (которые успешно сходились) не имела статистически значимых фиксированных эффектов (рис. S1B и S1C).
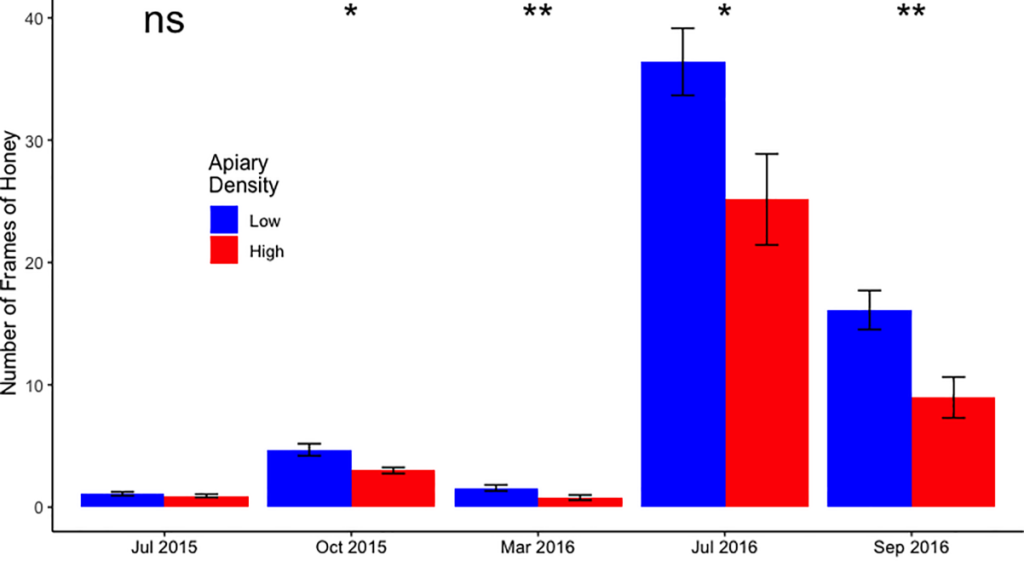
Тест Вилкоксона был применен к сравнению конфигурации пасеки в каждом месяце. Для подтверждения значимости множественных сравнений была выполнена процедура Бенджамина-Хохберга. Столбики ошибок представляют собой стандартную ошибку среднего значения. *: P<0.05; **: P<0,01; ns: несущественно.
Уровни клещей
GEE-модель данных по липким доскам показала значительное (P = 0,0188) положительное влияние инокуляции на уровень клещей, но не влияние конфигурации пасеки (рис. 3 и S1D рис.). Модель GEE не показала значительной связи между плотностью пасеки или инокуляцией на клещей на 100 расплодных клеток. Однако было обнаружено значительное взаимодействие (P = 0,0176) между статусом инокуляции и конфигурацией пасеки. Это взаимодействие указывает на то, что в расплоде было значительно больше клещей, когда колонии были одновременно инокулированы и располагались в конфигурации HD (рис. 4 и рис. S1E). GEE для уровней клещей, оцененных по спиртовым смывам, не содержал значимых членов (S1F рис.).

Обратите внимание, что очищенные колонии при обеих плотностях обработки оставались на низком уровне в течение первой зимы, в то время как инокулированные колонии постоянно увеличивались. Однако к концу первого года очищенные колонии достигли того же уровня заражения, что и инокулированные колонии. Столбики ошибок представляют собой стандартную ошибку среднего значения. GEE выявил значительный положительный эффект инокуляции на численность клещей (P = 0,0188).

A. Клещи в 100 ячейках расплода в зависимости от статуса инокуляции и конфигурации пасеки. B. GEE выявил значительное взаимодействие (P = 0,0176) между статусом инокуляции и конфигурацией. Планки ошибок на обоих графиках представляют собой стандартную ошибку среднего значения.
Анализ выживаемости
Анализ выживаемости по методу Кокса не выявил существенной связи между конфигурацией пасеки и статусом инокуляции или их взаимодействием (рис. 5A). Однако биномиальный анализ ошибок GLMM показал, что конфигурация пасеки была значительно связана с выживаемостью в зимний период (P = 0,037), причем колонии в конфигурации с меньшей плотностью имели больше шансов пережить зиму (рис. 5B).

A. Кривые выживаемости в зависимости от конфигурации пасеки и статуса инокуляции. Серые полосы показывают зимние месяцы — декабрь и январь. Обратите внимание на значительное снижение выживаемости во вторую зиму в конфигурации HD. B. Влияние плотности на зимнюю выживаемость (Примечание: колония, выжившая до начала второй зимы, учитывается на этом рисунке дважды). Логистическая модель зимней выживаемости показала, что колонии в конфигурациях с низкой плотностью имели значительно больше шансов пережить зиму (P = 0,037). Столбики ошибок на рисунке B показывают биномиальные доверительные интервалы.
Анализ дрейфа
Пчелы в конфигурации HD (N = 134 отслеженных пчел) значительно (P = 0,048) чаще дрейфовали на основе модели смешанных эффектов (рис. 6). 25,0% всех помеченных пчел в конфигурации HD дрейфовали, в то время как в конфигурации LD (N = 89 отслеженных пчел) дрейфовали только 7,5%. В четырех случаях мы наблюдали, как дрейфующая пчела входила в одну и ту же (неродовую) колонию несколько раз, и один раз мы обнаружили пчелу, которая вернулась в свою родную колонию. Почти весь дрейф в конфигурации HD был направлен на ближайшего соседа (1 м) к родной колонии фокальной пчелы (24 из 26 случаев дрейфа), и по одному случаю дрейфа на второго и шестого ближайшего соседа. В конфигурациях LD все три случая дрейфа произошли в ближайших соседних колониях (на расстоянии 10 м). С большой вероятностью (P = 0,047, коррелограмма Мантеля) дрейфующие пчелы попадали в колонию, ближайшую к их родной колонии, дрейф в колонии, расположенные дальше, не был значительным. Статус инокуляции клещей не был существенно связан с поведением дрейфующих пчел.
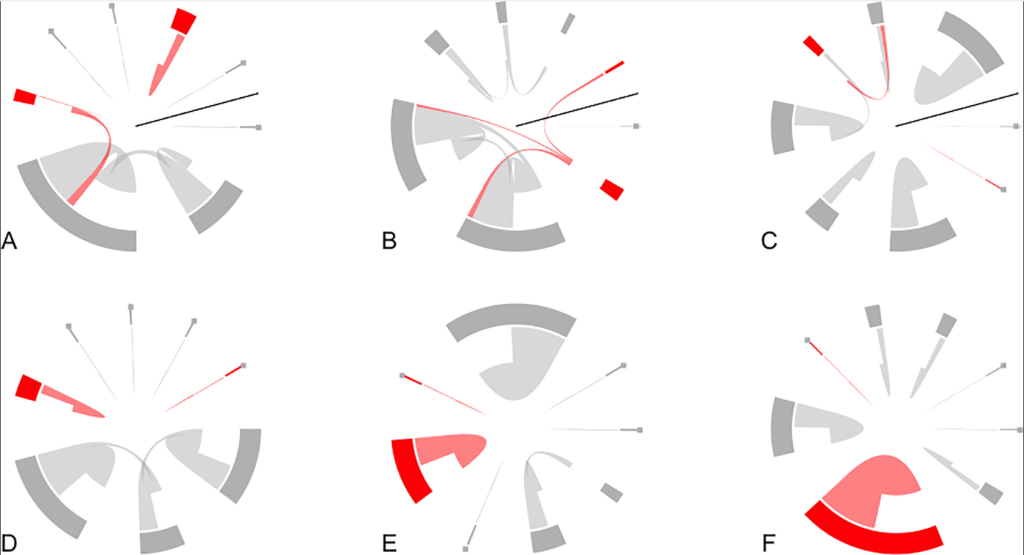
На каждой панели показаны восемь колоний как подсекции окружности круга (ширина этих подсечений указывает на количество уникальных пчел, наблюдаемых в колонии) и перемещения пчел (отдельные пчелы показаны тонкими линиями, так что большое количество наблюдаемых перемещений приводит к большим «клинообразным» формам). Направление дрейфа указывается тем, насколько близко линия движения отдельной пчелы находится к изображению колонии на окружности круга: пчела дрейфует к колонии, где линия находится близко к колонии, и выходит из колонии, где между линией и колонией большой разрыв. Красный цвет указывает на инокулированные колонии, а серый — на очищенные колонии. Панели A-C представляют конфигурации HD; они представлены здесь в круговом формате для сравнения с конфигурациями LD; черные линии в A-C представляют места, где линейное их расположение должно быть разделено, чтобы снова выстроиться в прямую линию. Панели D-F представляют дрейф в конфигурациях LD. Все смещения в D-F относятся к ближайшему соседу. Напротив, в A-C наблюдается повышенный дрейф, и есть два случая дрейфа за пределы ближайшего соседа (оба в B). Статус прививки колонии не оказал существенного влияния на дрейф.
Обсуждение
Обзор
Управляемые колонии медоносных пчел содержатся при плотности, которая значительно превышает естественную плотность в одичавших колониях. Современная практика пчеловодства, предпочитающая такую высокую плотность по логистическим причинам, по прогнозам, будет иметь пагубные последствия для распространения болезней и выживания колоний [26]. В ходе настоящей работы мы получили четыре основных результата, связанных с тем, как конфигурация пасеки влияет на здоровье медоносных пчел. Во-первых, конфигурация HD значительно снизила производство меда. Во-вторых, взаимодействие инокуляции и конфигурации HD привело к значительному повышению уровня заражения расплода медоносных пчел V. destructor. В-третьих, конфигурация HD оказала значительное негативное влияние на выживаемость колонии в зимний период. Наконец, при конфигурациях HD значительно увеличивался дрейф.
Сила колонии
Мы обнаружили, что производство меда снижалось в конфигурациях с высокой плотностью (рис. 2). Медовое скопление колонии в целом положительно связано с популяцией взрослых пчел [46], но поскольку мы не обнаружили сопутствующего сокращения ни популяции взрослых пчел, ни расплода, наши результаты лучше интерпретировать как прямой эффект либо 1) конфигурации плотности пасеки, либо 2) увеличения численности клещей V. destructor.
Во-первых, колонии медоносных пчел могут работать менее эффективно в конфигурации HD из-за путаницы или смешанных сигналов, что приводит к снижению эффективности фуражиров. Например, в одном исследовании было обнаружено, что скопление кормовых пчел побуждает кормовых пчел подавать сигналы другим пчелам о прекращении кормежки [20].
Во-вторых, увеличение численности клещей может негативно сказаться на запасах меда в колонии. Однако Мурилхас [47] не смог обнаружить прогрессирующего негативного воздействия клещей на расплод, взрослое население или запасы меда, пока численность клещей не достигла экстремально высокого уровня. Более того, он не смог обнаружить прямого влияния уровня клещей в колонии на запасы меда на одну пчелу в день и пришел к выводу, что предполагаемое воздействие клещей на запасы меда в колонии является косвенным эффектом разрушения популяций взрослых пчел. Наши данные указывают на увеличение количества клещей в расплоде инокулированных колоний в конфигурациях HD, но, как и в случае с Murilhas (2002), это произошло в конце эксперимента. Однако, в отличие от Murilhas (2002), запасы меда в наших инокулированных колониях конфигурации HD были стабильно ниже на протяжении большей части эксперимента. Короче говоря, влияние V. destructor на накопление меда, как на индивидуальном уровне, так и на уровне колонии, остается неоднозначным. Однако хорошо известно, что клещи V. destructor, помимо того, что сами вызывают патологию, передают патогенные вирусы [41,48], и вполне возможно, что наблюдаемое сокращение запасов меда было частично обусловлено этими вирусами. Важно отметить, что вирусы могут влиять на производительность колонии, воздействуя на поведение маток и плодовитость [49]. Для изучения этой возможности потребуются дальнейшие эксперименты.
Уровни клещей
Исходя из первоначальной инокуляции 400 клещей (2 колонии х 200 клещей), мы предсказывали, что уровень V. destructor в инокулированных и неинокулированных колониях на пасеках с высокой плотностью/визуально простой конфигурацией будет расти быстрее и оставаться более высоким на протяжении всего эксперимента по сравнению с низкой плотностью/визуально сложной конфигурацией. Этого не произошло, несмотря на то, что предыдущие полевые работы подтверждали эти ожидания. Например, одно исследование показало увеличение скорости инвазии и уровня популяции V. destructor в колониях, которые были окружены колониями с более высокой плотностью [50]. Другое исследование показало значительно меньшее количество клещей на пасеках, где колонии находились на расстоянии 100 м друг от друга, по сравнению с пасеками, где расстояние между колониями составляло 0 м или 10 м [51]. А генетические анализы свидетельствуют о значительной горизонтальной передаче клещей между колониями в условиях интенсивного пчеловодства [52].
Тем не менее, мы обнаружили значительное взаимодействие между статусом инокуляции и конфигурацией пасеки в уровне клещей в расплоде (рис. 4B), так что самые высокие уровни клещей наблюдались в инокулированных колониях в конфигурации HD. Этот результат может быть обусловлен временной задержкой, необходимой для увеличения количества клещей до высокого уровня, даже если колонии на пасеках с более высокой плотностью имели более высокий уровень передачи клещей по сравнению с пасеками с низкой плотностью.
В нашем исследовании инокуляция зарождающейся популяции клещей, будь то в условиях HD или LD, вызвала повышение уровня клещей в колонии, которое сохранялось, по крайней мере, пять месяцев, что является обратным отражением результатов Делаплана и Худа [33], которые показали сезонную пользу от ранней (февральской) обработки колоний, выходящих из зимы. Однако только на пасеках HD инокуляция также привела к увеличению доли клещей в расплоде. Поскольку только клещи в расплоде размножаются, процент клещей в расплоде является положительным показателем плодовитости популяции клещей [53]. Непонятно, как плотность пасеки могла повлиять на относительное распределение клещей между взрослыми особями и расплодом, особенно в нашем случае, когда площадь расплода не различалась по эффектам. Интересно сравнить эти результаты с результатами Нолана и Делаплана [51], которые, используя другой метод изменения конфигурации пасеки, показали более высокую численность клещей в конце сезона на пасеках с высокой плотностью после зарождающейся прививки 300 клещей, но не смогли показать различий в клещах в расплоде. Таким образом, хотя ландшафтные регуляторы, по-видимому, имеют значение для плодовитости и размера популяции V. destructor, для выяснения механизмов необходима дальнейшая работа.
Анализ выживаемости
Интересно, что анализ выживаемости в течение всего эксперимента не показал большей смертности инокулированных колоний по сравнению с очищенными колониями или различий между конфигурациями HD и LD. Отчасти это могло быть связано с другими факторами. Например, фоновая смертность, вызванная выбытием маток или другими факторами, могла скрыть влияние инфекции и плотности на смертность на ранних стадиях эксперимента. Кроме того, как описано выше, динамика клещей предполагает межколониальную передачу клещей, в результате чего очищенные и инокулированные колонии со временем становятся более равными по численности клещей.
Однако, когда анализ выживаемости был ограничен смертностью во время зимовки, мы обнаружили значительное влияние конфигурации пасеки. Действительно, в то время как на пасеках LD две зимы пережили ровно 50 процентов инокулированных колоний, на пасеках HD две зимы не пережила ни одна инокулированная колония (рис. 5A). То, что выживаемость в зимний период была значительно выше на пасеках с меньшей плотностью, является важным выводом для пчеловодов. Зимняя смертность в настоящее время является одной из самых больших проблем, с которыми сталкиваются пчеловоды в умеренных и холодных регионах земного шара [42,54,55], и наш эксперимент показывает, что эта смертность может быть значительно снижена путем управления колониями в конфигурации LD. Существует, по крайней мере, два фактора, которые могли способствовать повышению выживаемости в нашем эксперименте. Во-первых, конфигурации LD имели более высокое производство меда. Поскольку колонии в регионах с умеренным климатом нуждаются в достаточном количестве меда, чтобы пережить зиму [56], следует, что увеличение производства меда могло сыграть свою роль в повышении выживаемости. Хотя колонии в нашем исследовании кормили дополнительным сахарным сиропом (стандартизированным для всех колоний), что является стандартной практикой в управлении пчеловодством, могли существовать популяционные различия между колониями в их способности преобразовывать его в запасы меда [46], наряду с различиями в скорости сбора цветов, а также различиями в скорости потребления, которые мы не измеряли. Во-вторых, значительно более высокие уровни клещей в расплоде в инокулированных колониях в конфигурации HD в дату выборки перед второй зимой (рис. 4A), возможно, способствовали 100-процентной смертности оставшихся колоний в ту зиму.
Наши результаты согласуются с результатами другого исследования, в котором также было установлено, что выживаемость в течение зимы была значительно ниже на переполненной пасеке по сравнению с группой рассредоточенных колоний [5]. Однако в этом исследовании роение пчел не подавлялось, что делает эту работу потенциально менее применимой к обычной практике пчеловодства.
Дрейф
Давно известно, что расстояние, направление входа и расположение пасеки могут влиять на поведение медоносных пчел в дрейфе [57]. Мы использовали эти знания для минимизации дрейфа в наших конфигурациях LD, размещая колонии на расстоянии 10 м друг от друга по кругу, на разной высоте, с колониями, обращенными наружу, окрашенными в разные цвета и помеченными разными символами. Эти меры оказались эффективными, снизив дрейф с 25% на пасеке HD до 7,5% на пасеке LD. Эта количественная оценка важна для менеджеров, рассматривающих способы замедления передачи болезни, и для специалистов по моделированию болезней, работающих над параметризацией влияния пространства на распространение болезни. Эти наблюдаемые показатели дрейфа попадают в большие диапазоны доли дрейфующих особей (0-89%), обнаруженные в других исследованиях [29,58]. Наше наблюдение за четырьмя пчелами, оставшимися в неродовых колониях после дрейфа, позволяет предположить, что дрейфующие пчелы могут постоянно менять колонии после дрейфа, а не ходить туда-сюда, хотя это очень малые размеры выборки. Наш эксперимент также показал, что большинство дрейфов происходило между ближайшими соседями. В совокупности эти результаты свидетельствуют об относительно меньших объемах передачи болезни по сравнению с альтернативными вариантами, когда пчелы дрейфуют туда-сюда между родными и неродными колониями, или пчелы случайно залетают в любую колонию, что приводит к распространению клещей на большие расстояния от места первоначальной инокуляции. Хотя в нашем эксперименте не было возможности напрямую количественно оценить передачу клещей между колониями через дрейф, весьма вероятно, что увеличение скорости дрейфа увеличивает передачу болезни. Видеослежение за дрейфующими пчелами, которое мы использовали здесь [38], представляет собой плодотворный метод для дальнейшего изучения роли дрейфа в передаче болезни.
Заключение
Современное управление пасеками использует колонии в конфигурациях, подобных нашей процедуре HD (близко расположенные, визуально похожие колонии в линейных массивах на одной высоте) по практическим и логистическим причинам. Однако наш эксперимент с использованием воспроизведенных конфигураций HD и LD показал, что типичная практика управления (наша конфигурация HD) может быть вредной для здоровья и продуктивности колоний. Наши результаты показывают, что, снизив плотность пасеки и сделав колонии визуально отличимыми, пчеловоды могут повысить продуктивность колоний, снизить зимнюю смертность и потенциально уменьшить распространение болезней внутри пасеки за счет снижения дрейфа. Эти меры относительно скромны и могут быть реализованы во многих пчеловодческих хозяйствах.
Вспомогательная информация
Благодарности
Мы благодарны Н. Уивер, Б. Нолану, Н. Бич, Дж. Гаррисону, Б. Раусу и В. Джонсу за содержание и сбор измерений в колониях. Мы также благодарны Э. Доббсу, К. Беллу, К. Айерсу, З. Линчу, И. Хиньосе, А. Майрану, Э. Димиту, А. Мерчант, Э. Харрису, К. Бриссе, С. Тиггесу и Дж. Фаулеру за мечение пчел. Мы благодарим А. Мейрана, Э. Дымита, Н. Китли, Б. Мех, Х. Лейпоу, Х. Бока, А. Монгу, А. Файфа, Н. Кокса, С. Тиггеса, Л. Гилберт-Одем, К. Александера, Х. Наира, Д. Обиаго, Э. Резника, Х. Брэдшоу и О. Хендлера за подсчет смывов спирта и А. Джонсона за помощь в видеоанализе.
Исследования, представленные в данной публикации, были поддержаны Национальным институтом здравоохранения (R01-109501, для JCdR, BJB и KSD) и Национальным научным фондом (DGE-1444932, для TLD). Ответственность за содержание несут исключительно авторы, и оно не обязательно отражает официальную точку зрения Национальных институтов здравоохранения или Национального научного фонда.
Ссылки
1.vanEngelsdorp D, Meixner MD. A historical review of managed honey bee populations in Europe and the United States and the factors that may affect them. J Invertebr Pathol. 2010;103:S80–95. pmid:19909973
View Article PubMed/NCBI Google Scholar
2.Potts SG, Ngo HT, Biesmeijer JC, Breeze TD, Dicks LV, Garibaldi LA, et al. The Assessment Report on Pollinators, Pollination and Food Production of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services. Bonn, Germany: Secretariat of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; 2017 p. 556.
3.Ratnieks FLW A. Piery M, Cuadriello I. The natural nest and nest density of the Africanized honey bee (Hymenoptera, Apidae) near Tapachula, Chiapas, Mexico. Can Entomol. 1991;123:353–9.
4.Seeley T. Honey bees of the Arnot Forest: A population of feral colonies persisting with Varroa destructor in the northeastern United States. Apidologie. 2007;38:19–29.
5.Seeley TD, Smith ML. Crowding honeybee colonies in apiaries can increase their vulnerability to the deadly ectoparasite Varroa destructor. Apidologie. 2015;46:716–27.
6.Malthus TR. An essay on the principle of population as it affects the future improvement of society, with remarks on the speculations of Mr. Godwin, M. Condorcet, and other writers. Lond J Johns. 1798;
7.Stiling P. Density-dependent processes and key factors in insect populations. J Anim Ecol. 1988;57:581–93.
8.Lizaso JLS, Goñi R, Reñones O, Charton JAG, Galzin R, Bayle JT, et al. Density dependence in marine protected populations: a review. Environ Conserv. 2000;27:144–58.
9.Grace JB. The factors controlling species density in herbaceous plant communities: An assessment. Perspect Plant Ecol Evol Syst. 1999;2:1–28.
10.Fowler CW. A review of density dependence in populations of large mammals. Curr Mammal. Springer, Boston, MA: Genoways H.H. (eds); 1987. p. 401–41.
11.Bowers MA, Matter SF. Landscape ecology of mammals: Relationships between density and patch size. J Mammal. 1997;78:999–1013.
12.Oaten A, Murdoch WW. Functional response and stability in predator-prey systems. Am Nat. 1975;109:289–98.
13.Gordon DM, Kulig AW. Founding, foraging, and fighting: Colony size and the spatial distribution of Harvester Ant nests. Ecology. 1996;77:2393–409.
14.Cao TT, Dornhaus A. Ants under crowded conditions consume more energy. Biol Lett. 2008;4:613. pmid:18765354
View Article PubMed/NCBI Google Scholar
15.Yarahmadi P, Miandare HK, Fayaz S, Caipang CMA. Increased stocking density causes changes in expression of selected stress- and immune-related genes, humoral innate immune parameters and stress responses of rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2016;48:43–53. pmid:26549176
View Article PubMed/NCBI Google Scholar
16.Huzzey JM, DeVries TJ, Valois P, von Keyserlingk MAG. Stocking density and feed barrier design affect the feeding and social behavior of dairy cattle. J Dairy Sci. 2006;89:126–33. pmid:16357274
View Article PubMed/NCBI Google Scholar
17.Harbo JR. Worker-bee crowding affects brood production, honey production, and longevity of honey bees (Hymenoptera: Apidae). J Econ Entomol. 1993;v. 86:1672–8.
18.Rueppell O, Kaftanouglu O, Page RE. Honey bee (Apis mellifera) workers live longer in small than in large colonies. Exp Gerontol. 2009;44:447–52. pmid:19389467
View Article PubMed/NCBI Google Scholar
19.Thom C. The tremble dance of honey bees can be caused by hive-external foraging experience. J Exp Biol. 2003;206:2111–6. pmid:12771160
View Article PubMed/NCBI Google Scholar
20.Lau CW, Nieh JC. Honey bee stop-signal production: temporal distribution and effect of feeder crowding. Apidologie. 2010;41:87–95.
21.Neumann P, Moritz RFA, van Praagh J. Queen mating frequency in different types of honey bee mating apiaries. J Apic Res. 1999;38:11–8.
22.Anderson RM, May RM, Anderson B. Infectious Diseases of Humans: Dynamics and Control. Reprint edition. Oxford; New York: Oxford University Press; 1992.
23.Hudson PJ, Rizzoli A, Grenfell BT, Heesterbeek H, Dobson AP, editors. The Ecology of Wildlife Diseases. 1 edition. New York: Oxford University Press; 2002.
24.Ramsey D, Spencer N, Caley P, Efford M, Hansen K, Lam M, et al. The effects of reducing population density on contact rates between brushtail possums: Implications for transmission of bovine tuberculosis. J Appl Ecol. 2002;39:806–18.
25.Lively CM. The ecology of virulence. Ecol Lett. 2006;9:1089–95. pmid:16972872
View Article PubMed/NCBI Google Scholar
26.Brosi BJ, Delaplane KS, Boots M, de Roode JC. Ecological and evolutionary approaches to managing honeybee disease. Nat Ecol Evol. 2017;1:1250–62. pmid:29046562
View Article PubMed/NCBI Google Scholar
27.Rosenkranz P, Aumeier P, Ziegelmann B. Biology and control of Varroa destructor. J Invertebr Pathol. 2010;103, Supplement:S96–119. pmid:19909970
View Article PubMed/NCBI Google Scholar
28.Kralj J, Fuchs S. Parasitic Varroa destructor mites influence flight duration and homing ability of infested Apis mellifera foragers. Apidologie. 2006;37:577–87.
29.Free JB. The drifting of honey-bees. J Agric Sci. 1958;51:294–306.
30.Aliano N, Ellis MD. A strategy for using powdered sugar to reduce Varroa populations in honey bee colonies. J Apic Res. 2005;44:54–7.
31.Milani N. Activity of oxalic and citric acids on the mite Varroa destructor in laboratory assays. Apidologie. 2001;32:127–38.
32.Traynor KS, Rennich K, Forsgren E, Rose R, Pettis J, Kunkel G, et al. Multiyear survey targeting disease incidence in US honey bees. Apidologie. 2016;47:325–47.
33.Delaplane KS, Hood WM. Economic threshold for Varroa jacobsoni Oud. in the southeastern USA. Ap. 1999;30:383–95.
34.Delaplane KS, van der Steen J, Guzman-Novoa E. Standard methods for estimating strength parameters of Apis mellifera colonies. J Apic Res. 2013;52:1–12.
35.Fries I, Aarhus A, Hansen H, Korpela S. Comparison of diagnostic methods for detection of low infestation levels of Varroa jacobsoni in honey-bee (Apis mellifera) colonies. Exp Appl Acarol. 1991;10:279–87.
36.Branco MR, Kidd NAC, Pickard RS. A comparative evaluation of sampling methods for Varroa destructor (Acari: Varroidae) population estimation. Apidologie. 2006;37:452–61.
37.Delaplane KS, Hood WM. Effects of delayed acaricide treatment in honey bee colonies parasitized by Varroa jacobsoni and a late-season treatment threshold for the south-eastern USA. J Apic Res. 1997;36:125–32.
38.Rossetti BJ, Dynes T, Brosi B, de Roode JC, Kong J. GRAPHITE: A graphical environment for scalable in situ video tracking of moving insects. Methods Ecol Evol. 2017;00:1–9.
39.Højsgaard S, Halekoh U, Yan J. The R Package geepack for Generalized Estimating Equations. J Stat Softw. 2006;15:1–11.
40.R Core Team. R: A language and environment for statistical computing [Internet]. Vienna, Austria: R Foundation for Statistical Computing; 2017. Available from: http://www.R-project.org/
41.Dainat B, Evans JD, Chen YP, Gauthier L, Neumann P. Dead or alive: Deformed Wing Virus and Varroa destructor reduce the life span of winter honeybees. Appl Environ Microbiol. 2012;78:981–7. pmid:22179240
View Article PubMed/NCBI Google Scholar
42.van Dooremalen C, Gerritsen L, Cornelissen B, van der Steen JJM, van Langevelde F, Blacquière T. Winter survival of individual honey bees and honey bee colonies depends on level of Varroa destructor infestation. PLOS ONE. 2012;7:e36285. pmid:22558421
View Article PubMed/NCBI Google Scholar
43.Rondeau V, Mazroui Y, Gonzalez JR. frailtypack: An R package for the analysis of correlated survival data with frailty models using penalized likelihood estimation or parametrical estimation. J Stat Softw. 2012;47:1–28.
44.Bates D, Maechler M, Bolker B, Walker S. Fitting linear mixed-effects models using lme4. J Stat Softw. 2015;67:1–48.
45.Oksanen J, Blanchet FG, Kindt R, Legendre P, Minchin PR, O’hara RB, et al. Package ‘vegan’ (version 2.5–2). Community Ecol Package Version [Internet]. 2018;2. Available from: https://CRAN.R-project.org/package=vegan
46.Farrar C. The influence of colony populations on honey production. J Agric Res. 1937;54:945–54.
47.Murilhas AM. Varroa destructor infestation impact on Apis mellifera carnica capped worker brood production, bee population and honey storage in a Mediterranean climate. Apidologie. 2002;33:271–81.
48.Nazzi F, Brown SP, Annoscia D, Del Piccolo F, Di Prisco G, Varricchio P, et al. Synergistic parasite-pathogen interactions mediated by host immunity can drive the collapse of honeybee colonies. PLoS Pathog. 2012;8:e1002735. pmid:22719246
View Article PubMed/NCBI Google Scholar
49.Fievet J, Tentcheva D, Gauthier L, de Miranda J, Cousserans F, Colin ME, et al. Localization of deformed wing virus infection in queen and drone Apis mellifera L. Virol J. 2006;3:16. pmid:16569216
View Article PubMed/NCBI Google Scholar
50.Frey E, Rosenkranz P. Autumn invasion rates of Varroa destructor (Mesostigmata: Varroidae) into honey bee (Hymenoptera: Apidae) colonies and the resulting increase in mite populations. J Econ Entomol. 2014;107:1–8.
51.Nolan MP, Delaplane KS. Distance between honey bee Apis mellifera colonies regulates populations of Varroa destructor at a landscape scale. Apidologie. 2017;48:8–16.
52.Dynes T, de Roode JC, Lyons JI, Berry JA, Delaplane KS, Brosi BJ. Fine scale population genetic structure of Varroa destructor, an ectoparasitic mite of the honey bee (Apis mellifera). Parasitology. 2017;48:93–101.
53.Harbo J R, Harris J W. Selecting honey bees for resistance to Varroa jacobsoni. Apidologie. 1999;30:183–96.
54.vanEngelsdorp D, Hayes J, Underwood R, Pettis J. A survey of honey bee colony losses in the United States, fall 2008 to spring 2009. J Apic Res. 2010;49:14.
55.Dainat B, Evans JD, Chen YP, Gauthier L, Neumann P. Predictive markers of honey bee colony collapse. PLoS ONE. 2012;7:e32151. pmid:22384162
View Article PubMed/NCBI Google Scholar
56.Free JB, Racey PA. The effect of the size of honeybee colonies on food consumption, brood rearing and the longevity of the bees during winter. Entomol Exp Appl. 1968;11:241–9.
57.Jay SC. Drifting of honeybees in commercial apiaries. III. Effect of apiary layout. J Apic Res. 1966;5:137–48.
58.Neumann P, Moritz RFA, Mautz D. Colony evaluation is not affected by drifting of drone and worker honeybees (Apis mellifera L.) at a performance testing apiary. Apidologie. 2000;31:67–79.
Добавить комментарий