
Аннотация
Современные методы селекционной селекции демонстрируют высокий потенциал для улучшения экономически важных признаков медоносных пчел. Однако при селекции могут игнорироваться фундаментальные правила естественного отбора. Целью данного исследования был анализ влияния отбора и местной адаптации на некоторые важные селекционные признаки Apis mellifera carnica. Мы сравнили открыто спаренных маток трех различных происхождений: местные хорватские генотипы, которые находятся под контролируемым отбором в течение пяти поколений (MS), местный генотип, который никогда не подвергался селекции (NS), и, наконец, неместный генотип из долгосрочной немецкой селекционной программы (HS). В целом, колонии с королевами из селекционных программ имели лучшие показатели по оборонительному поведению, спокойствию и роению. Напротив, колонии из группы НС показали более высокую выраженность признаков устойчивости к Varroa destructor. Хотя группа HS показала самые высокие результаты по большинству поведенческих признаков, низкий индекс зимовки колоний из этой группы после обоих периодов зимовки указывает на отсутствие местной адаптации. Данное исследование подчеркивает актуальность селекционной работы для повышения устойчивости популяций медоносных пчел и достижения местной адаптации.
1 Введение
Селекционная селекция медоносных пчел (Apis mellifera) имеет давние традиции во многих европейских странах, где она практикуется различными способами и в разной степени (Lodesani and Costa 2003; Hatjina et al. 2014a). Сегодня селекция пчел преимущественно направлена на улучшение признаков, представляющих интерес для пчеловодства и оказывающих важное влияние на пчеловодческую отрасль, таких как снижение оборонительного поведения, увеличение производства меда или снижение склонности к роению (Ruttner 1972; Möbus 1983; Büchler et al. 2013; Tiesler et al. 2016; Uzunov et al. 2017). Преимущества селекции хорошо известны и могут сильно повлиять на экономический успех пчеловодства. Например, на основе программы селекции с использованием оценки селекционной ценности, производство меда в немецкой популяции Carnica увеличилось на 0,7 кг в год с 1996 года (Bienefeld 2016), что означает ежегодное увеличение стоимости продукции примерно на 3 миллиона евро только для немецких пчеловодов.
Однако нынешние усилия по селекционной селекции медоносных пчел также подвергаются сомнению, поскольку иногда они могут игнорировать важные признаки жизненной истории (такие как воспроизводство или выживание) и противодействовать механизмам естественного отбора, которые могут способствовать повышению устойчивости (Neumann and Blacquière 2017; Blacquière and Panziera 2018). Как следствие, можно ожидать отрицательной реакции отбора для признаков, которые отбираются напрямую, но коррелируют неблагоприятно. Например, отбор на высокую продуктивность расплода и низкую склонность к роению, с целью достижения более высокого производства меда, может снизить социальный иммунитет колонии (Meunier 2015) и ее естественную устойчивость к патогенам. В естественной среде обитания медоносные пчелы заселяют сравнительно небольшие гнездовые полости, что приводит к частому и раннему роению и меньшим колониям. Как следствие, в таких колониях меньше проблем с эктопаразитическим клещом Varroa destructor (Fries and Bommarco 2007; Loftus et al. 2016). С другой стороны, современное пчеловодство с эффективным контролем V. destructor минимизирует давление естественного отбора на пчел для развития механизмов резистентности (Fries et al. 2006) и установления сбалансированных отношений хозяина и паразита.
Однако может существовать и компромисс между признаками истории жизни (такими как размножение) и параметрами здоровья (Webster and Woolhouse 1999; Schwenke et al. 2016). Эванс и Петтис (2005) сообщают, что колонии с пчелами, устойчивыми к парше, вызываемой личинками Paenibacillus, показали более низкую продуктивность по сравнению с более восприимчивыми колониями, что свидетельствует о возможном компромиссе между устойчивостью и продуктивностью. Другой пример компромисса между устойчивостью и воспроизводством дает естественная выжившая популяция в Швеции, где колонии производят меньше расплода как часть адаптации к паразитам (Fries and Bommarco 2007). Аналогичные наблюдения были зарегистрированы для устойчивых к варроа «приморских пчел» в Германии (Rosenkranz et al. 2010) и США (Tarpy et al. 2007), где эти пчелы производят меньше расплода и меда по сравнению с местным поголовьем без резистентности, но, похоже, более устойчивы к клещам. В отличие от этого, компромисс между поведенческими признаками (такими как защитное поведение или склонность к роению) и урожайностью меда кажется менее значимым, поскольку положительная генетическая корреляция этих признаков была показана в нескольких исследовательских отчетах (Bienefeld and Pirchner 1991; Brascamp et al. 2016). Отбор на усиление гигиенического поведения, по-видимому, не оказывает негативного влияния на продуктивность (Spivak and Reuter 1998) или индивидуальный врожденный иммунитет пчел (Harpur et al. 2014).
В последнее время влияние взаимодействия генотипа и окружающей среды на поведение, продуктивность и выживаемость пчел было хорошо признано и изучено, что привело к общей рекомендации использовать адаптированных к местным условиям пчел (Costa et al. 2012a; Büchler et al. 2014; Hatjina et al. 2014b; Uzunov et al. 2014). Хотя в целом не наблюдалось разницы в нагрузке патогенов между колониями местных и неместных генотипов (Meixner et al. 2014), ожидается более высокая склонность к заболеваниям у пчел, завезенных из разных регионов (Francis et al. 2014). Кроме того, механизмы устойчивости к болезням, которые колонии могут проявлять в одной среде, могут быть не выражены в другой среде (Corrêa-Marques et al. 2002; Seeley 2007).
Целью данного исследования было сравнить поведение, продуктивность и устойчивость к V. destructor у различных генотипов A. m. carnica при сравнении местных и неместных генотипов и различных режимов отбора. В частности, мы сравнили два местных генотипа, которые либо (1) никогда не находились под искусственным отбором, либо (2) произошли от селекционной программы, с (3) интродуцированным и высокоселекционным генотипом. Полученные результаты обсуждаются с точки зрения преимуществ селекции, ее влияния на устойчивость пчел и воздействия местной адаптации.
2 Материалы и методы
2.1 Территория исследования и управление колониями
Исследование проводилось в Осечко-Бараньском уезде, Чеминац, Хорватия (N 45°40′12″; E 18°40′40″; высота над уровнем моря 90 м) с октября 2015 года по февраль 2017 года. Всего было включено 60 колоний трех различных карниольских генотипов в стандартных ульях Лангстрота. Все колонии были созданы в начале мая 2015 года с четырьмя сотами с расплодом, занятыми пчелами, двумя сотами с пыльцой и медом и спаренной королевой. В июле все существующие королевы в экспериментальных колониях были заменены на подопытных королев. В августе 2015 года все колонии были обработаны полосками CheckMite® для снижения и выравнивания уровня заражения V. destructor. После этого обработка конкретных колоний проводилась в индивидуальном порядке каждый раз, когда уровень заражения взрослой популяции пчел достигал 10%. Проверки колоний начались 5 октября 2015 года, когда большинство рабочих пчел в колониях представляли собой дочерей подопытных маток (Büchler et al. 2013).
2.2 Генетическое происхождение групп
Первый генотип (N = 13, без селекционных усилий, группа NS) происходил из популяции, которая, насколько нам известно, никогда не подвергалась селекционной селекции. Королевы этой группы происходили из изолированного района в центральной части Хорватии, где большинство пчеловодов все еще держат пчел в традиционных ульях-лежаках. Опрос, проведенный среди пчеловодов, подтвердил, что в этом районе нет ни миграционного пчеловодства, ни торговли роями, ни маток. Королевы из этой группы были сводными сестрами и спаривались в пределах района происхождения. Второй генотип (N = 35, средняя селекционная нагрузка, группа MS) получен из местной популяции в северо-восточной Хорватии, отобранной по основным пчеловодческим признакам (защитное поведение, спокойствие, роение, медопродуктивность) в течение пяти поколений. Тестовые королевы происходили от трех маток, которые были сводными сестрами и спаривались на брачной станции, насыщенной местными трутнями другого генотипа, но происходящими из той же программы разведения. Королевы третьего генотипа (N = 12, высокая селекционная нагрузка, > 10 поколений, группа HS) были полусестрами, происходящими от одной королевы-селекционера (DE-7-45-73-2013, см. родословную на сайте www.beebreed.eu) из высокоселекционной популяции Carnica (по продуктивности, мягкости, спокойствию, низкому роению, гигиеническому поведению и низкому распространению клещей) Института пчеловодства в Кирхгайне (Германия) и открыто спаривались с местными трутнями в Риеке (Хорватия).
Различия в селекционной работе между МС и ГС заключаются в (1) количестве селекционируемых поколений, (2) способе оценки племенной ценности и (3) контроле спаривания маток. Все поколения маток МС были открыто спарены на неизолированной станции спаривания, насыщенной трутнями, выведенными из отобранных линий, в то время как спаривание маток маток ГС поколений осуществлялось на изолированной островной станции спаривания с полным контролем происхождения трутней.
2.3 Проверенные параметры
Все исследуемые параметры перечислены в таблице I, вместе с номером и датой измерения и использованными методами. Индекс зимовки (ИЗ) рассчитывался как соотношение между количеством пчел при первом весеннем осмотре после зимы и количеством пчел при последнем осеннем осмотре перед зимой. Первый период зимовки (измеряемый от начала значительного снижения активности пчел до первого более крупного поступления пыльцы) длился 163 дня, а второй — 122 дня. Количество чистого добытого меда для каждой колонии рассчитывали путем измерения веса отдельных ящиков с медом до и после добычи. Образцы взрослых пчел для определения степени заражения V. destructor брали из медовых суперов или из верхнего расплода в сентябре и октябре (Costa et al. 2012b; Büchler et al. 2013). Хотя колонии регулярно проверялись на зараженность клещами, для дальнейшего анализа использовались только те даты отбора проб (даты с 1 по 7 в таблице I), когда во всех колониях были сопоставимые условия для развития клещей (колонии не обрабатывались с июля 2015 года до конца июля 2016 года).
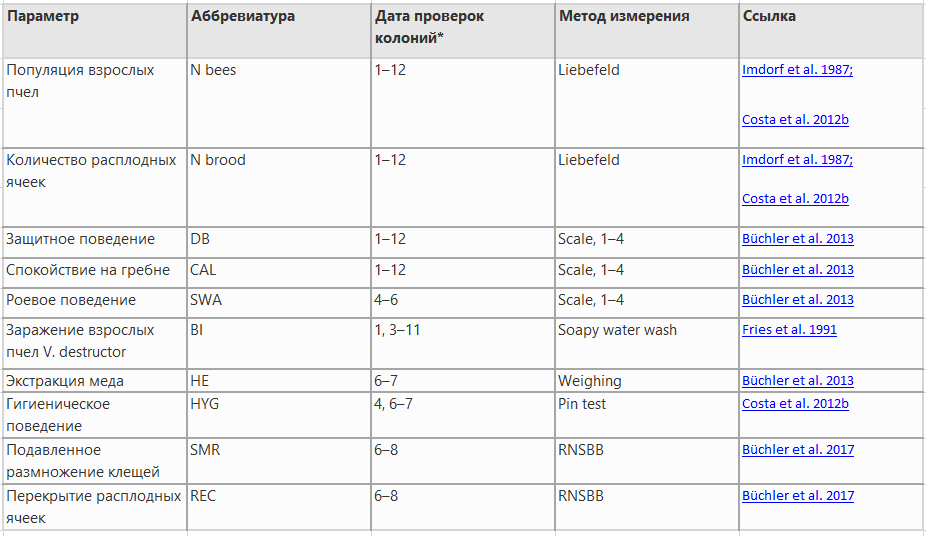
Анализ доли клещей V. destructor, которые не смогли размножиться (SMR), а также раскупоривание и повторное закупоривание выводка (REC) определяли в соответствии с протоколом RNSBB (Büchler et al. 2017). Вкратце, образцы расплода с куколками на стадии развития между 7 и 12 днями после укупорки вскрывали до тех пор, пока не обнаруживали 35 или по крайней мере 10 клеток, зараженных одиночными клещами-находками. Клещ-найденыш считался нерепродуктивным (1), если он не произвел никакого потомства, (2) если не было потомства мужского пола или исключительно мужского пола, или (3) если старшая дочь была слишком молода, чтобы достичь взрослой стадии и спариться до закрытия пчелы. Образцы расплода для анализа собирали один раз в месяц в течение июня, июля и августа.
2.4 Статистический анализ
При анализе всех исследуемых параметров использовалась общая линейная модель (ОЛМ). Генотип и год или месяц, а также взаимодействие между ними считались фиксированными эффектами. Различия между группами определялись с помощью теста Бонферрони (Bonferroni post hoc analysis test). Все значения представлены как скорректированные средние величины (LS means ± SE). Если колония погибала, в статистическом анализе использовались все данные, собранные до этого события. Все статистические анализы проводились с использованием программного обеспечения STATISTICA 10 (StatSoft 2011).
3 Результаты
3.1 Сила колоний
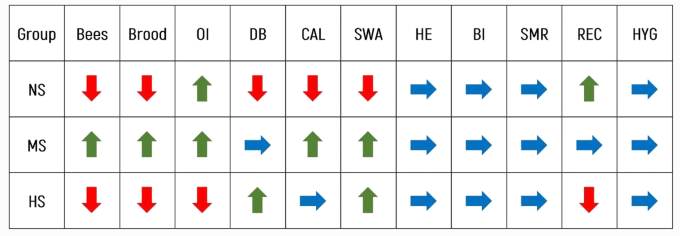
GLM-анализ показал, что генотип и месяц как фиксированные факторы имели значительное (p < 0,01) влияние на количество пчел, в то время как не было эффекта (p = 0,448) от взаимодействия месяц × генотип (Дополнение к таблице I). Группа МС имела значительно большее количество пчел по сравнению с группами НС и ГС (Таблица II). Анализ также показал значительное влияние месяца (p < 0,01) и генотипа (p < 0,05) на количество расплодных ячеек (Дополнительная таблица I). Взаимодействие Месяц × Генотип не оказало влияния (p = 0,153) на количество расплода. Как и в случае с количеством пчел, группа МС имела наибольшее количество расплодных ячеек (Таблица II). С середины марта до конца апреля количество пчел (среднее среднее фенотипическое значение всех колоний) увеличилось с 6275 до 16 667, а количество расплодных ячеек — с 6338 до 26 700. Наибольшее количество пчел (38 942) было зарегистрировано в колонии из группы НС в июне, а наибольшее количество расплодных ячеек (36 960) за время исследования было зарегистрировано в конце апреля в колонии из группы МС.

3.2 Оборонительное поведение, спокойствие на гребне и склонность к роению
Защитное поведение значительно (p < 0,01) зависело от генотипа и месяца (дополнительная таблица II). Самый высокий балл, т.е. наименее оборонительные пчелы, были зарегистрированы в группе HS, которая значительно отличалась (p < 0,01) от других экспериментальных групп (Таблица II). Генотип, месяц и их взаимодействие также показали значительный эффект (p < 0,01) на спокойствие (Дополнение к таблице II). Самый высокий балл был зарегистрирован в группе МС, а самый низкий — в группе НС, которая имела значительно более низкий балл (p < 0,01), чем две другие группы (Таблица II). На склонность к роению влиял генотип (Дополнительная таблица III), и группа NS имела значительно (p < 0,01) более низкий балл, т.е. более высокую склонность к роению, по сравнению с двумя другими группами (Таблица II).
3.3 Производство меда
В связи с неблагоприятными климатическими условиями во время основного потока нектара, в 2016 году в тестовых колониях было достигнуто необычно низкое производство меда. Среднее количество добытого меда составило 10,9 кг. Это очень мало по сравнению со средним многолетним показателем производства меда на колонию в Хорватии, который находится в диапазоне от 18,2 кг для пчеловодов-любителей до 41,4 кг для профессиональных пчеловодов (Штефанич и др. 2004). Наибольшее количество меда было добыто из группы ГС, а наименьшее — из группы МС, но эти различия не были значительными (Таблица II, Дополнительная таблица V).
3.4 Выживаемость и способность к зимовке
Все колонии пережили первый зимний период. Однако после второй зимы были зарегистрированы различные потери колоний среди групп: 75% колоний НС выжили, 70,4% из группы МС и 90% из группы ГС. На индекс зимовки (ИЗ) значительно повлиял (p < 0,01) генотип, но не год (Дополнительная таблица IV). Только одна колония с уровнем заражения V. destructor более 8% взрослых пчел в предыдущем году пережила зиму с более чем 6000 пчел. Общее скорректированное среднее значение ОИ после первой зимы составило 75,38 ± 2,61%, а после второй зимы — 66,15 ± 3,82%. Местные колонии МС и НС имели почти одинаковый ОИ, значительно более высокий, чем колонии из группы ГС (Таблица III).

3.5 Зараженность V. destructor и признаки устойчивости
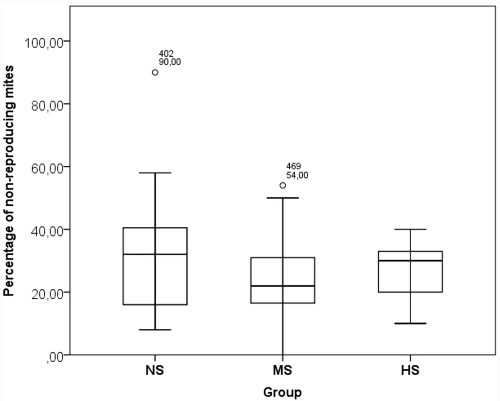
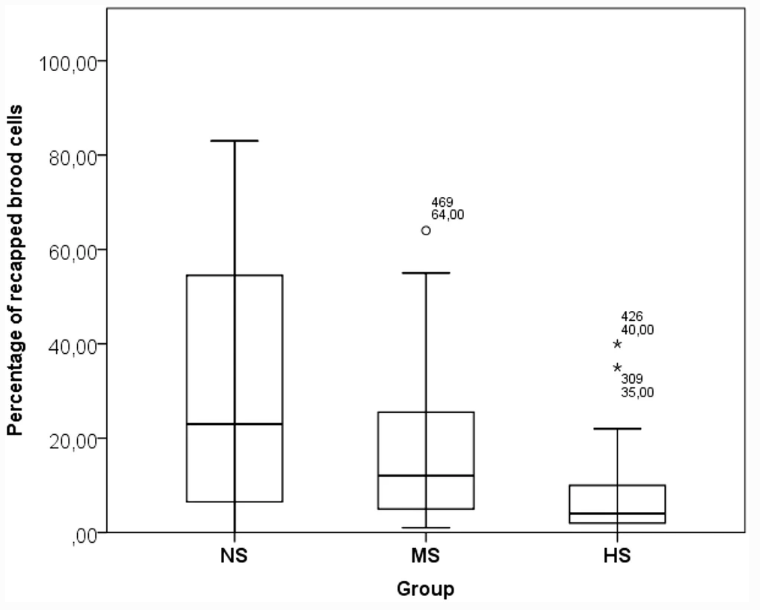
Анализ GLM для зараженности взрослых пчел клещами показал значительный эффект Месяца как фиксированного фактора (Дополнительная таблица III). Значительных различий в уровне зараженности взрослых пчел между группами не наблюдалось (Таблица III). Хотя не было существенной разницы в гигиеническом удалении куколок из расплодных ячеек (Таблица III, Дополнительная таблица VII), самая высокая тенденция была отмечена в группе ГС. Что касается ВСР, то не было выявлено влияния ни одного из факторов в модели (Дополнительная таблица VI). Наибольшая доля нерепродуцирующих клещей была зарегистрирована в группе NS, но она существенно не отличалась от двух других групп (Таблица III). Факторы Генотип и Месяц оказали значительное влияние на повторное заклещевывание выводковых ячеек (Дополнительная таблица VI). Значительные различия были зарегистрированы между группами, с самой высокой активностью по укупорке в группе NS и самой низкой в HS (Таблица III). Кроме того, в группе NS была обнаружена самая высокая вариабельность SMR и REC (рис. 1 и 2).
4 Обсуждение
Результаты нашего эксперимента ясно показывают преимущества долгосрочной селекции и разведения на поведение пчел, особенно для признаков оборонительного поведения, спокойствия на гребне и склонности к роению. Показатели по этим трем признакам были значительно выше в группах, которые возникли в результате селекционных программ. В сравнительном исследовании поведенческих признаков 16 различных генотипов (Узунов и др. 2014), одним из самых высокооцененных оказался A. m. carnica, происходящий из Кирхгайна, что соответствует генотипу HS в данном исследовании (Таблица IV). В то же время эти авторы обнаружили, что колонии, возглавляемые местными царицами, были в среднем менее оборонительными, чем колонии с неместными царицами, таким образом, предполагая, что взаимодействие генотип-среда оказывает значительное влияние на этот признак. Однако в данном исследовании мы не смогли подтвердить это наблюдение. Напротив, нелокальная группа HS демонстрировала наименее оборонительное поведение, указывая на то, что генетический эффект отбора может преобладать над потенциальным негативным эффектом для этого признака из-за изменения окружающей среды. Как спокойствие на гребне, так и оборонительное поведение значительно зависели от периода года, и самые низкие показатели были зарегистрированы в конце зимы (февраль и март) и осенью (октябрь). Это может быть влиянием погодных условий в это время года или следствием возрастной структуры пчел, когда в колонии находятся в основном старшие пчелы.
Высокая склонность к роению была одним из основных признаков, наблюдаемых в группе NS, и здесь можно отметить явное влияние генетического происхождения пчел. Королевы из группы НС происходили из традиционных ульев-лежаков, которые предлагают ограниченное пространство для развития и, таким образом, способствуют более раннему и частому роению (Simpson and Riedel 1963). Хотя ранней весной группа HS, из-за низкого индекса зимовки, была слабее двух других групп по количеству пчел и расплодных ячеек, в сезон роения не было зарегистрировано значительной разницы в силе колоний. Более того, эта группа имела тенденцию быть самой сильной во время второго пика роения в конце июня. Следовательно, различия в силе колоний, вероятно, не могут быть причиной меньшей выраженности роевого поведения в группе HS.
Тем не менее, различия в силе колоний между группами наблюдались. Если рассматривать развитие колоний, способность к зимовке и производство меда как показатели местной адаптации, то у неместного поголовья ГС можно наблюдать явные негативные последствия изменения окружающей среды.
От медоносных пчел на коммерческих пасеках сегодня ожидают выживания и высокой продуктивности в различных экологических условиях по всему миру. Желательные признаки лишь нескольких экономически предпочтительных подвидов повлияли на распространение этих коммерчески успешных запасов в ущерб местным медоносным пчелам (Ruttner 1988; Moritz et al. 2005). Однако, не в последнюю очередь вызванная дискуссиями о здоровье медоносных пчел и потерях колоний в последние десятилетия, местная адаптация сегодня все больше признается как один из важных факторов, влияющих на поведение, продуктивность и выживание пчел (Costa et al. 2012a; Büchler et al. 2014; Hatjina et al. 2014b; Uzunov et al. 2014). Заметно, что в нашем эксперименте колонии, возглавляемые неместными матками (группа HS), достигали пика развития примерно на 1 месяц позже, чем местные группы NS и MS. Это может свидетельствовать о дезадаптации к местным экологическим условиям в нашем районе исследований, где период цветения растений начинается на несколько недель раньше, чем в Германии, откуда прибыли эти королевы. Аналогичное явление наблюдали Дражич и др. (2014), которые сообщили, что в Хорватии колонии, возглавляемые матками из Австрии, выводили меньше расплода весной по сравнению с местными колониями. Значительно меньшее количество пчел и расплодных ячеек, наблюдаемое в группе NS, может быть следствием ее происхождения. Традиционные ульи-лежаки имеют небольшое и ограниченное пространство, что не позволяет увеличить общий потенциал колонии. С другой стороны, матки из группы МС содержались в более просторных ульях и систематически отбирались для роста колонии, и преимущества отбора можно увидеть в наших результатах. Другим важным показателем дезадаптации является успешность зимовки, которая измерялась путем расчета OI. Хотя фактор года не повлиял на ОИ (хотя две зимы были очень разными по продолжительности и интенсивности), между генотипами наблюдались значительные различия, в основном за счет низкого ОИ, зарегистрированного в группе HS. Поскольку во время последнего осеннего осмотра колоний в обоих сезонах не было зафиксировано существенной разницы в силе колоний, очевидно, на перезимовку влияли другие факторы. Заражение колоний V. destructor сокращает продолжительность жизни зимующих пчел (Kovac and Crailsheim 1988; van Dooremalen et al. 2012; Steinmann et al. 2015). Однако это не может объяснить низкую ОИ в группе ГС, поскольку между группами не было разницы в зараженности клещами. Даже если основные механизмы не могут быть идентифицированы по нашим результатам, мы предполагаем, что более низкий ОИ неместной группы ГС, зарегистрированный после обеих зим, может указывать на отсутствие адаптации. Местные пчелы адаптируются к местным климатическим условиям не только поведением, т.е. развитием расплода в течение года (Strange et al. 2007; Dražić et al. 2014), но и на метаболическом уровне (Parker et al. 2010). Кроме того, местные пчелы легче справляются с местными патогенами (Francis et al. 2014; Hatjina et al. 2014b; Salmela et al. 2015). Отсутствие адаптации также могло повлиять на производство меда в группе ГС, которая показала не столь высокие результаты и произвела меньше меда, чем ожидалось, возможно, из-за более низкой степени эксплуатации ресурсов окружающей среды. Вместе с крайне плохими погодными условиями, зарегистрированными в период цветения Brassica napus и Robinia pseudoacacia (21 дождливый день, зарегистрированный с 15 апреля по 15 мая), это могло повлиять на потребление нектара во время этих двух основных медоносов в районе исследований. Следовательно, первый сбор меда произошел только относительно поздно, после цветения Tilia sp. и Helianthus annuus. Тем не менее, возможно, что этот неполный сезон скрыл потенциальную разницу в способности производства меда между группами.
Гигиеническое поведение является признаком с высокой наследуемостью (Rothenbuhler 1964; Pérez-Sato et al. 2009; Facchini et al. 2019), и ожидалось, что в группе ГС оно будет выражено значительно сильнее, чем в двух других группах. Тот факт, что эти различия не были значительными, может быть следствием открытого спаривания маток из группы HS с неселекционированными трутнями неизвестного происхождения, что подчеркивает отцовское влияние на наследование этого признака (Pérez-Sato et al. 2009).
Несмотря на отсутствие существенной разницы между группами в отношении подавления размножения V. destructor, в группе НС была отмечена удивительно высокая доля нерепродуктивных клещей (> 30%). Это может быть следствием укупорки выводка, которая была наиболее выражена в этой группе. Было показано, что вскрытие и повторное закрытие выводковых ячеек играет ключевую роль в популяциях, устойчивых к клещам (Oddie et al. 2018; Martin et al. 2019). Популяция, из которой произошли королевы NS, подвергается обработке против клещей в значительно меньшей степени (один раз в год в октябре). Это, вероятно, способствует усилению давления отбора в сторону устойчивости к V. destructor, например, из-за более высокой приспособленности трутней из менее восприимчивых колоний в условиях естественного спаривания (Büchler et al. 2006). Поэтому более высокое поведение этой группы может указывать на то, что эта популяция могла выработать определенную степень устойчивости к клещу. Тот факт, что эти различия по одному из значимых признаков устойчивости не коррелируют с различиями в зараженности пчел клещами, можно объяснить как следствие дрейфа, поскольку колонии на исследовательской пасеке располагались близко друг к другу рядами (Jay 1966; Seeley and Smith 2015). Очевидно, что более высокая изменчивость REC в группе NS по сравнению с группой HS указывает на большую возможность селекции на этот признак в популяции NS.
В настоящее время необходимость разведения и содержания жизнестойких пчел обозначена как наиболее перспективное устойчивое решение (Büchler et al. 2010; Uzunov et al. 2017) для преодоления проблемы клеща Варроа. Экономические затраты на обработку колоний и потери колоний высоки (Neumann and Carreck 2010; Brodschneider et al. 2018), а химические обработки, хотя и теряют свою значимость из-за устойчивости клещей к различным компонентам (Martin 2004; Sammataro et al. 2005), могут загрязнять продукты пчеловодства (Богданов 2006). Один из возможных путей достижения устойчивого пчеловодства в присутствии клещей — позволить коэволюцию пчел и V. destructor с как можно меньшим вмешательством, но соблюдая пороговые уровни ущерба от заражения клещами (Liebig 2001; Currie and Gatien 2006; Genersch et al. 2010). Если исходить из более «приближенного к природе» подхода и поддерживать достаточно высокий уровень заражения V. destructor в период роения/ спаривания, то трутни из наиболее приспособленных колоний будут иметь больше шансов спариться и передать свои гены, и, следовательно, будут способствовать повышению жизнестойкости будущих поколений. В конечном итоге, выявление механизмов, лежащих в основе устойчивости к клещам, может быть использовано в селекционных программах для содействия процессу отбора. Однако это необходимо делать в региональном масштабе, принимая во внимание адаптацию местных пчел к конкретной среде, в которой они обитают.
В заключение, колонии, возглавляемые королевами, которые происходят из селекционных программ, показали значительно лучшие показатели по основным коммерчески признанным поведенческим признакам. С другой стороны, колонии из группы без селекции показали более высокую выраженность признаков устойчивости (SMR и REC), что указывает на возможное улучшение этих признаков в результате естественного отбора и местной адаптации. Кроме того, низкий индекс перезимовки и более медленное весеннее развитие указывают на отсутствие адаптации у группы HS. Данное исследование предоставляет дополнительные доказательства важности разведения пчел по аспектам устойчивости и значимости местной адаптации для производительности колоний.
Электронный дополнительный материал
Ссылки
1.Bienefeld, K. (2016) Breeding success or genetic diversity in honey bees? Bee World 93(11), 40-44. Google Scholar
2.Bienefeld, K., Pirchner, F. (1991) Genetic correlations among several colony characters in the honey bee (Hymenoptera: Apidae) taking queen and worker effects into account. Ann. Entomol. Soc. Am. 84(3), 324–331. Google Scholar
3.Blacquière, T., Panziera, D. (2018) A plea for use of honey bees’ natural resilience in beekeeping. Bee World 95(2), 34-38. Google Scholar
4.Bogdanov, S. (2006) Contaminants of bee products. Apidologie 37(1), 1-18. CAS Google Scholar
5.Brascamp, E.W., William, A., Boigenzahn, C., Bijma, P., Veerkamp, R.F. (2016) Heritabilities and genetic correlations for honey yield, gentleness, calmness and swarming behaviour in Austrian honey bees. Apidologie 47(6), 739-748. Google Scholar
6.Brodschneider, R., Gray, A., Adjlane, N., Ballis, A., Brusbardis, V. et al. (2018) Multi-country loss rates of honey bee colonies during winter 2016/2017 from the COLOSS survey. J. Apic. Res. 57(3), 452-457. Google Scholar
7.Büchler, R., Moritz, R., Garrido, C., Bienefeld, K., Ehrhardt, K. (2006) Male fitness in relation to colony development and varroosis infection. Proceedings of the 2nd European Conference of Apidology, 10.14.09, Prague, pp. 44–45.
8.Büchler, R., Berg, S., Le Conte, Y. (2010) Breeding for resistance to Varroa destructor in Europe. Apidologie 41(3), 393-408.
Google Scholar
9.Büchler, R., Andonov, S., Bienefeld, K., Costa, C., Hatjina, F. et al. (2013) Standard methods for rearing and selection of Apis mellifera queens. J. Apic. Res. 52(1), 1–30. Google Scholar
10.Büchler, R., Costa, C., Hatjina, F., Andonov, S., Meixner et al. (2014) The influence of genetic origin and its interaction with environmental effects on the survival of Apis mellifera L. colonies in Europe. J. Apic. Res. 53(2), 205–214. Google Scholar
11.Büchler, R., Costa, C., Mondet, F., Kezić, N., Kovačić, M. (2017) Screening for low Varroa mite reproduction (SMR) and recapping in European honey bees. Available at:
https://dev.rescol.org/rnsbbweb/wp-content/uploads/2017/11/RNSBB_SMR-recapping_protocol_2017_09_11.pdf (accessed on 2 February 2018.)
12.Corrêa-Marques, M. H., De Jong, D., Rosenkranz, P., Gonçalves, L. S. (2002) Varroa-tolerant Italian honey bees introduced from Brazil were not more efficient in defending themselves against the mite Varroa destructor than Carniolan bees in Germany. Genet. Mol. Res. 1(2), 153–158.PubMed Google Scholar
13.Costa, C., Lodesani, M., Bienefeld, K. (2012a) Differences in colony phenotypes across different origins and locations: evidence for genotype by environment interactions in the Italian honeybee (Apis mellifera ligustica)? Apidologie 43(6), 634–642. Google Scholar
14.Costa, C., Büchler, R., Berg, S., Bienkowska, M., Bouga, M., et al. (2012b) A Europe-wide experiment for assessing the impact of genotype–environment interactions on the vitality and performance of honey bee colonies: experimental design and trait evaluation. J. Apic. Sci. 56(1), 147–158. Google Scholar
15.Currie R.W., Gatien P. (2006) Timing acaricide treatments to prevent Varroa destructor (Acari: Varroidae) from causing economic damage to honey bee colonies. Can. Entomol. 138(2), 238–252. Google Scholar
16.Dražić, M. M., Filipi, J., Prđun, S., Bubalo, D., Špehar, M. et al. (2014) Colony development of two Carniolan genotypes (Apis mellifera carnica) in relation to environment. J. Apic. Res. 53(2), 261-268. Google Scholar
17.Evans J.D., Pettis J.S. (2005) Colony-level impacts of immune responsiveness in honey bees, Apis mellifera. Evolution 59(10), 2270-2274. CAS PubMed Google Scholar
18.Facchini, E., Bijma, P., Pagnacco, G., Rizzi, R., Brascamp, E.W. (2019) Hygienic behaviour in honeybees: a comparison of two recording methods and estimation of genetic parameters. Apidologie 50(2), 163-172. Google Scholar
19.Francis, R.M., Amiri, E, Meixner, M., Kryger, P, Gajda, A. et al. (2014) Effect of genotype and environment on parasite and pathogen levels in one apiary—a case study. J. Apic. Res. 53(2), 230-232. Google Scholar
20.Fries, I., Bommarco, R. (2007) Possible host–parasite adaptations in honey bees infested by Varroa destructor mites. Apidologie 38(6), 525–533. Google Scholar
21.Fries, I., Aarhus, A., Hansen, H., Korpela, S. (1991) Comparison of diagnostic methods for detection of low infestation levels of Varroa jacobsoni in honey-bee (Apis mellifera) colonies. Exp. Appl. Acarol. 10(3-4), 279–287. Google Scholar
22.Fries, I., Imdorf, A., Rosenkranz, P. (2006) Survival of mite infested (Varroa destructor) honey bee (Apis mellifera) colonies in a Nordic climate. Apidologie 37(5), 564–570. Google Scholar
23.Genersch, E. von der Ohe, W., Kaatz, H., Schroeder, A., Otten, C., Büchler, R., Berg, S., Ritter, W., Mühlen, W., Gisder, S., Meixner, M., Liebig, G., Rosenkranz, P. (2010) The German bee monitoring project: a long term study to understand periodically high winter losses of honey bee colonies. Apidologie 41(3), 332–352. CAS Google Scholar
24.Harpur, B.A., Chernyshova, A., Soltani, A., Tsvetkov, N., Mahjoorighasrodashti, M., Xu, Z., et al. (2014) No genetic tradeoffs between hygienic behaviour and individual innate immunity in the honey bee, Apis mellifera. PLoS ONE 9(8), e104214.
PubMed PubMed Central Google Scholar
25.Hatjina, F., Bienkowska, M., Charistos, L., Chlebo, R., Costa, C., et al. (2014a) A review of methods used in some European countries for assessing the quality of honey bee queens through their physical characters and the performance of their colonies. J. Apic. Res. 53(3), 337-363. Google Scholar
26.Hatjina, F., Costa, C., Büchler, R., Uzunov, A., Drazic, M. et al. (2014b) Population dynamics of European honey bee genotypes under different environmental conditions. J. Apic. Res. 53(2), 233-247. Google Scholar
27.Imdorf, A., Buehlmann, G., Gerig, L., Kilchenmann, V., Wille, H. (1987) Überprüfung der Schätzmethode zur Ermittlung der Brutfläche und der Anzahl Arbeiterinnen in freifliegenden Bienenvölkern. Apidologie, 18(2), 137-146.
28.Jay, S.C. (1966) Drifting of honeybees in commercial apiaries. II. Effect of various factors when hives are arranged in rows. J. Apic. Res. 5(2), 103-111. Google Scholar
29.Kovac, H., Crailsheim, K. (1988) Lifespan of Apis mellifera carnica Pollm. infested by Varroa jacobsoni Oud. in relation to season and extent of infestation. J. Apic. Res. 27(4), 230-238. Google Scholar
30.Liebig, G. (2001) How many varroa mites can be tolerated by a honey bee colony? Apidologie 32(5), 482– 484. Google Scholar
31.Lodesani, M., Costa, C. (2003) Bee breeding and genetics in Europe. Bee World, 84(2), 69-85. Google Scholar
32.Loftus, J.C., Smith, M.L., Seeley, T.D. (2016) How honey bee colonies survive in the wild: testing the importance of small nests and frequent swarming. PLoS ONE 11(3), e0150362. PubMed PubMed Central Google Scholar
33.Martin, S.J. (2004) Acaricide (pyrethroid) resistance in Varroa destructor. Bee World 85(4), 67-69. Google Scholar
34.Martin, S.J., Hawkins, G.P., Brettell, L.E., Reece, N., Correia-Oliveira et al. (2019) Varroa destructor reproduction and cell re-capping in mite-resistant Apis mellifera populations. Apidologie, 1-14.
35.Meixner, M. D., Francis, R. M., Gajda, A., Kryger, P., Andonov, S. et al. (2014) Occurrence of parasites and pathogens in honey bee colonies used in a European genotype–environment interactions experiment. J. Apic. Res. 53(2), 215–219. Google Scholar
36.Meunier, J. (2015) Social immunity and the evolution of group living in insects. Phil. Trans. R. Soc. B 370(1669), 20140102. PubMed Google Scholar
37.Möbus, B. (1983) Bee breeding in Germany. In: B., Möbus, B; J., van Praagh (Eds). Pedigree bee breeding in western Europe. British Isles Bee Breeders Association; Derby, UK. pp. 6–12
38.Moritz, R. F. A., Härtel, S., Neumann, P. (2005) Global invasion of western honeybee (Apis mellifera) and the consequences for biodiversity. Écoscience, 12(3), 289–301. Google Scholar
39.Neumann, P., Blacquière, T. (2017) The Darwin cure for apiculture? Natural selection and managed honeybee health. Evol. Appl. 10(3), 226–230. PubMed Google Scholar
40.Neumann, P., Carreck, N. (2010) Honey bee colony losses. J. Apic. Res. 49(1), 1. Google Scholar
41.Oddie, M., Büchler, R., Dahle, B., Kovačić, M., Le Conte, Y. et al. (2018). Rapid parallel evolution overcomes global honey bee parasite. Scientific reports 8, 7704. PubMed PubMed Central Google Scholar
42.Parker, R., Melathopoulos, A.P., White, R., Pernal, S.F., Guarna, M.M., et al. (2010) Ecological adaptation of diverse honey bee (Apis mellifera) populations. PLoS ONE 5(6), e11096. PubMed PubMed Central Google Scholar
43.Pérez-Sato, J. A., Châline, N., Martin, S. J., Hughes, W. O. H., & Ratnieks, F. L. W. (2009) Multi-level selection for hygienic behaviour in honeybees. Heredity, 102(6), 609–615. PubMed Google Scholar
44.Rosenkranz, P., Aumeier, P., Ziegelmann, B. (2010) Biology and control of Varroa destructor. J. Invertebr. Pathol. 103(Suppl 1), S96-119. PubMed Google Scholar
45.Rothenbuhler, W.C. (1964) Behavior genetics of nest cleaning in honey bees. Responses of F1 and backcross generations to disease-killed brood. Am Zool. 4(2), 111–123. CAS PubMed Google Scholar
46.Ruttner, H. (1972) Technical recommendations for methods of evaluating performance of bee colonies. In: F. Ruttner (Ed), Controlled mating and selection of the honey bee. Apimondia, Bucharest, Romania. pp. 87-92.
47.Ruttner, F. (1988) Biogeography and taxonomy of honeybees. Springer Verlag, Berlin. Google Scholar
48.Salmela, H., Amdam, G.V., Freitak, D. (2015) Transfer of immunity from mother to offspring is mediated via egg-yolk protein vitellogenin. PLoS Pathog. 11(7), e1005015. PubMed PubMed Central Google Scholar
49.Sammataro, D., Untalan, P., Guerro, F., Finley, J. (2005) The resistance of Varroa mites (Acari: Varroidae) to acaricides and the presence of esterase. Internat. J. Acarol. 31(1), 67-74. Google Scholar
50.Schwenke, R.A., Lazzaro, B.P., Wolfner, M.F. (2016) Reproduction–immunity trade-offs in insects. Annu. Rev. Entomol. 61(1), 1-18.
Google Scholar
51.Seeley, T. (2007) Honey bees of the Arnot Forest: a population of feral colonies persisting with Varroa destructor in the northeastern United States. Apidologie 38(1), 19–29. Google Scholar
52.Seeley, T.D., Smith, M.L. (2015) Crowding honeybee colonies in apiaries can increase their vulnerability to the deadly ectoparasite Varroa destructor. Apidologie 46(6), 716-727. Google Scholar
53.Simpson, J., Riedel, I.B.M. (1963) The factor that causes swarming in honeybee colonies in small hives. J. Apic. Res 2(1), 50–54.
Google Scholar
54.Spivak, M., Reuter, G. S. (1998) Performance of hygienic honey bee colonies in a commercial apiary. Apidologie 29(3), 291–302.
Google Scholar
55.StatSoft, Inc. (2011). STATISTICA (data analysis software system), version 10. www.statsoft.com.
56.Štefanić, I., Štefanić, E., Puškadija, Z., Kezić, N., Grgić, Z. (2004) Beekeeping in the Republic of Croatia. Bee World 85(1), 19–21.
Google Scholar
57.Steinmann, N., Corona, M., Neumann, P., Dainat, B. (2015) Overwintering is associated with reduced expression of immune genes and higher susceptibility to virus infection in honey bees. PLoS ONE, 10(6): e0129956.
PubMed PubMed Central Google Scholar
58.Strange, J.P., Garnery, L., Sheppard. W.S. (2007) Persistence of the Landes ecotype of Apis mellifera mellifera in southwest France: confirmation of a locally adaptive annual brood cycle trait. Apidologie, 38(3), 259-267.
59.Tarpy, D.R., Summers, J., Keller, J.J., Hensey, W. (2007) Comparing pairs of Russian and Italian colonies by new beekeepers in North Carolina. Am. Bee J. 147(2), 149–152. Google Scholar
60.Tiesler, F.K., Bienefeld, K., Büchler, R. (2016) Selektion bei der Honigbiene. Buschhausen Druck- und Verlagshaus, Herten.
Google Scholar
61.Uzunov, A., Costa, C., Panasiuk, B., Meixner, M., Kryger, P., et al. (2014) Swarming, defensive and hygienic behaviour in honey bee colonies of different genetic origin in a Pan-European experiment. J. Apic. Res. 53(2): 248–260. Google Scholar
62.Uzunov, A., Brascamp, E.W., Büchler, R. (2017) The basic concept of honey bee breeding programs. Bee World 94(3): 84-87.
Google Scholar
63.van Dooremalen, C., Gerritsen, L., Cornelissen, B., van der Steen, J.J.M., van Langevelde, F., et al. (2012) Winter survival of individual honey bees and honey bee colonies depends on level of Varroa destructor infestation. PLoS ONE 7(4): e36285.
PubMed PubMed Central Google Scholar
64.Webster, J.P., Woolhouse, M.E.J. (1999) Cost of resistance: relationship between reduced fertility and increased resistance in a snail–schistosome host–parasite system. Proc. R. Soc. Lond. B 266(1417), 391-396. Google Scholar
Добавить комментарий