
Аннотация
Колонии медоносных пчел (Apis mellifera) ценятся за предоставляемые ими услуги по опылению. Однако в некоторых странах, включая США, смертность колоний возросла до неустойчивого уровня. Преобразование ландшафта в монокультурное сельское хозяйство, вероятно, играет определенную роль в этой повышенной смертности, поскольку сокращаются источники пищи, доступные для медоносных пчел. Многие землевладельцы и организации в регионе Верхнего Среднего Запада США хотели бы восстановить/восстановить исконные места обитания прерий. С ростом осведомленности общественности о высокой смертности пчел, многие землевладельцы и пчеловоды задаются вопросом, могут ли эти восстановленные прерии значительно улучшить питание колоний медоносных пчел. Удобно, что у медоносных пчел есть уникальный коммуникационный сигнал, называемый «танец виляния», который указывает местоположение цветочных пятен, воспринимаемых фуражирами как высокодоходные источники пищи. Мы использовали эти коммуникационные сигналы, чтобы ответить на два основных вопроса: Во-первых, существует ли какая-либо часть сезона, в которую кормовая сила колонии медоносных пчел будет посвящать большую часть своих усилий по привлечению (танцы виляния) цветочным пятнам в прериях? Во-вторых, будут ли медоносные пчелы рекламировать определенные таксоны местных цветов прерий как выгодные источники пыльцы? Мы расшифровали 1528 танцев виляния в колониях, расположенных рядом с двумя большими реконструированными прериями. Мы также собрали пыльцу с подгруппы танцующих пчел, которую затем проанализировали, чтобы определить таксон рекламируемых цветов. Большинство пчел рекламировали цветочные пятна за пределами реконструированных прерий, но доля пчел, рекламирующих источники нектара в прериях, значительно увеличилась в конце лета/осенью на одном участке. Медоносные пчелы рекламировали семь местных таксонов прерий как выгодные источники пыльцы, хотя три наиболее часто рекламируемых таксона пыльцы были неместными. Наши результаты показывают, что включение некоторых местных таксонов цветов прерий в реконструированные прерии может увеличить шансы на то, что колонии будут использовать эти прерии в качестве основных источников пищи в период наибольшего роста колоний и производства меда
1. Введение
В последние десятилетия многие популяции насекомых во всем мире столкнулись с быстрым сокращением численности из-за вызванных человеком изменений ландшафта, включая массовое увеличение площади земель, отведенных под монокультурное сельское хозяйство [1,2,3,4]. Сокращение популяций пчел, в частности, вызвало тревогу, поскольку пчелы оказывают важнейшие услуги по опылению [5,6], способствуя развитию мировой экономики и улучшая питание людей за счет удешевления выращивания разнообразных фруктов и овощей [5]. Наиболее широко используемый вид опылителей сельскохозяйственных культур, Apis mellifera L., европейская медоносная пчела, ежегодно оказывает услуги по опылению только в США на сумму около 14 миллиардов долларов [7]. В последние годы пчеловоды отмечают повышенную смертность колоний медоносных пчел в нескольких регионах, включая США [8,9].
Хотя воздействие патогенов, паразитов и пестицидов, несомненно, способствует высокой смертности колоний, ключевую роль, вероятно, играет плохое питание [6,9,10,11,12,13,14]. Как и все виды пчел, медоносные пчелы нуждаются как в пыльце — основном источнике белков и липидов, так и в нектаре — основном источнике углеводов [15,16,17]. В регионах с умеренным климатом колониям необходимо собрать примерно 25 кг пыльцы [17,18] и потенциально более 300 кг нектара [19], чтобы функционировать летом и пережить холодную зиму. Помимо количества, очень важно и качество рациона. Разнообразные источники пыльцы помогают медоносным пчелам бороться с патогенами и паразитами [20,21,22,23] и повышают их способность к детоксикации пестицидов [24]. Колонии, живущие в умеренной зоне, должны реагировать на частую смену видов цветущих цветов с весны до осени [25,26] и могут переживать периоды безводья, когда температура остается высокой, но цветет мало цветов, приносящих пользу [27,28].
Многие группы по всему миру заинтересованы в том, чтобы помочь сохранить здоровые популяции пчел, высаживая цветы для пчел [29,30,31]. Одновременно многие организации в более широком смысле заинтересованы в восстановлении исконных мест обитания, которые были преобразованы для сельского хозяйства или других видов использования человеком [32]. До европейской колонизации верхний Средний Запад США в основном состоял из прерий, определяемых как луга умеренного климата с умеренным количеством осадков и глубоко укоренившимися многолетними травами [33,34]. Сегодня от первоначальных прерий осталось менее 2% [34], и существует большой интерес к восстановлению исконных мест обитания прерий [35]. К сожалению, правительства и организации, заинтересованные в помощи пчелам, часто имеют ограниченную информацию о том, как различные схемы управления землей [36] или смеси семян [29] повлияют на кормовой успех пчел. Проекты восстановления прерий, скорее всего, принесут пользу местным видам пчел, особенно пчелам, которые специализируются на цветах прерий [37,38,39]. Менее ясно, в какой степени неместные медоносные пчелы будут привлекаться и использовать участки цветов в восстановленных прериях.
Уникальный жизненный цикл и кормовая стратегия медоносных пчел могут повлиять на использование ими цветочных ресурсов в прериях. Медоносные пчелы являются универсальными кормилицами, которые имеют очень широкий кормовой диапазон, при этом большинство кормовых поездок происходит в пределах 4 км от гнезда [40,41,42,43], но некоторые поездки достигают 14 км [44,45]. Колонии медоносных пчел содержат тысячи фуражиров, которые могут общаться друг с другом о местонахождении наиболее выгодных участков цветов с помощью сигнала, называемого танцем виляния, который является уникальным для пчел рода Apis [26,44]. Этот сигнал включает повторяющиеся восьмерки, в которых танцовщица покачивает брюшком вперед-назад во время прямой средней части восьмерки, называемой виляющим бегом. При этом танцующие пчелы получают вектор, содержащий направление на цветочную грядку относительно азимута солнца и расстояние до нее [44]. Фуражиры будут танцевать только для того, чтобы объявить о расположении цветочных участков, которые они воспринимают как выгодные (с благоприятным соотношением полученных питательных веществ и затраченной энергии) [46]. Эти особенности кормового поведения медоносных пчел могут привести к тому, что колонии будут концентрироваться на самых плотных участках цветов и игнорировать более редкие цветы в не приносящих прибыль травах, как это обычно бывает в реконструированных прериях. Однако плотность и размер цветочных пятен в прериях меняются в течение сезона по мере цветения различных видов цветов. Даже если медоносные пчелы воспринимают цветы в прериях как выгодные ресурсы только в течение части кормового сезона, доступ к прериям может способствовать здоровью колонии, поставляя нектар или пыльцу в условиях недостатка непаровых источников пищи [47].
Танец виляния медоносной пчелы дает нам возможность понять, как фуражиры медоносной пчелы воспринимают ресурсы, которые они встречают в ландшафте вокруг своих ульев [36]. На уровне колонии доля танцев, рекламирующих цветочные пятна в данной среде обитания, служит мерой процесса принятия решений, который распределяет фуражиров между типами среды обитания на основе их относительной прибыльности [36]. На уровне особи, если фуражир приносит пищу с определенного вида цветов и танцует, рекламируя место, которое она посетила, это указывает на то, что она воспринимает эти цветы как достаточно выгодные для привлечения сородичей. В настоящее время возможно определить, какой цветочный таксон рекламировался в танце, по пыльце, которую приносил танцор, но не по нектару, который приносили танцоры [48,49]. Кроме того, характеристики танцев, рекламирующих источники нектара, могут дать более тонкую информацию о предполагаемой прибыльности ресурса. Общее количество виляний, которые танцовщица нектара совершает в танце, коррелирует с ее оценкой прибыльности рекламируемого ресурса [46]. Эта связь была многократно продемонстрирована с искусственными кормушками с сахаром и водой [50,51,52], но до сих пор не была продемонстрирована с пыльцой или заменителями пыльцы ([53], но см. [54]). Многочисленные исследования также показали, что колонии склонны рекламировать места на больших расстояниях, чтобы найти выгодные источники нектара в периоды дефицита [27,40,42,55].
Поэтому мы воспользовались информацией, содержащейся в танцах пчел, чтобы ответить на два основных вопроса о том, как колонии медоносных пчел воспринимают цветы в прериях: во-первых, существует ли какая-либо часть сезона, в которой кормовая сила колонии медоносных пчел посвящает большую часть своих усилий по привлечению (танцы) источникам пыльцы или нектара в прериях? Чтобы лучше понять сезонные изменения в доле танцев, рекламирующих прерии, мы задали два дополнительных вопроса: 1) В те времена года, когда большая часть нектарных танцев рекламирует источники в прериях, включают ли танцы для мест в прериях больше виляний и тем самым указывают на более высокую предполагаемую прибыльность, чем танцы для мест вне прерий, и 2) Есть ли сезонные изменения в среднем расстоянии от рекламируемых нектарных источников? Для нашего второго основного вопроса мы спросили, будут ли медоносные пчелы рекламировать участки с цветами местных прерий как высококачественные источники пыльцы, и если да, то какие таксоны они будут рекламировать? Чтобы ответить на эти вопросы, мы поместили колонии медоносных пчел в наблюдательные ульи со стеклянными стенками с доступом к двум большим реконструированным прериям. Ульи со стеклянными стенками позволили нам записывать танцы, исполняемые членами этих колоний в течение лета и начала осени. На основе этих записей мы расшифровали информацию о направлении и расстоянии в танцах, отобразили их в виде распределения плотности вероятности, используя подход Байесовского моделирования [56], и определили, какой процент танцев рекламировал места в прериях в разные части сезона. На участках с сезонным изменением доли танцев, рекламирующих источники пищи в прериях, мы также количественно оценили количество виляний в танце, чтобы определить, воспринимают ли пчелы цветы в прериях как более выгодные по сравнению с цветами за пределами прерий. Мы также исследовали, есть ли сезонные изменения в среднем расстоянии до рекламируемых источников нектара. Наконец, чтобы составить карту и определить таксоны, которые фуражиры считали выгодными источниками пыльцы, мы отловили часть танцующих пчел, которые несли груз пыльцы, и идентифицировали источник пыльцы с помощью микроскопии и ДНК-штрихкодирования.
2. Материалы и методы
2.1 Разрешения и согласования
Данное исследование не требовало никаких разрешений, поскольку мы не приносили в жертву ни одной пчелы и не собирали материал с цветов, внесенных в список находящихся под угрозой исчезновения штата Миннесота или федерального правительства США. Мы получили разрешение от Карлтонского колледжа на проведение исследований цветов вдоль определенных трансект через их реконструированные прерии Cowling Arboretum, а также получили разрешение от Belwin Conservancy на проведение исследований и сбор цветов в их реконструированных прериях.
2.2 Места проведения исследования
В рамках данного исследования мы разместили колонии на двух участках: Belwin Conservancy (http://www.belwin.org/) в Афтоне, штат Миннесота (UTM 15N: 516102 E, 4976403 N; широта/долгота: 44.940869° N, 92.795909° W) и возле дендрария Коулинг Карлтонского колледжа (https://apps.carleton.edu/campus/arb/) в Нортфилде, штат Миннесота (UTM 15N: 488949 E, 4923159 N; широта/долгота: 44.461653° N, 93.138922° W; рис. 1). Мы наблюдали колонии с 9 июня по 1 сентября в 2015 году и с 16 мая по 24 июня в 2016 году в Belwin Conservancy и с 4 июня по 4 сентября в 2017 году в Carleton College. Belwin Conservancy — некоммерческая организация, занимающаяся сохранением и восстановлением местных экосистем, включая 300 акров реконструированной высокотравной прерии, и мы разместили колонии между двумя крупнейшими непрерывными участками прерии (менее чем в 2 км от самых дальних краев всех прерий Belwin). В дендрарии Каулинг мы разместили колонии в доме напротив одного участка восстановленной высокотравной прерии, где они имели доступ к 160 акрам восстановленной прерии в радиусе 2 км. Оба участка были засажены местными прерийными травами и злаками (от 5 до 40 лет назад в зависимости от участка и участка земли) и управляются комбинацией периодического выжигания, выпаса бизонов (в Belwin Conservancy) и целенаправленного удаления некоторых инвазивных видов (например, Digitalis lanata в Belwin Conservancy). Мы нанесли на карту края прилегающих участков прерий, используя файлы форм ArcMap, предоставленные Belwin Conservancy (Н. Филлипс, личное сообщение) и Carleton College (В.-Х. Фу и Н. Бракер, личное сообщение), которые мы немного изменили на основе спутниковых снимков участков от ESRI (www.esri.com), используя ArcGIS Desktop [57]).
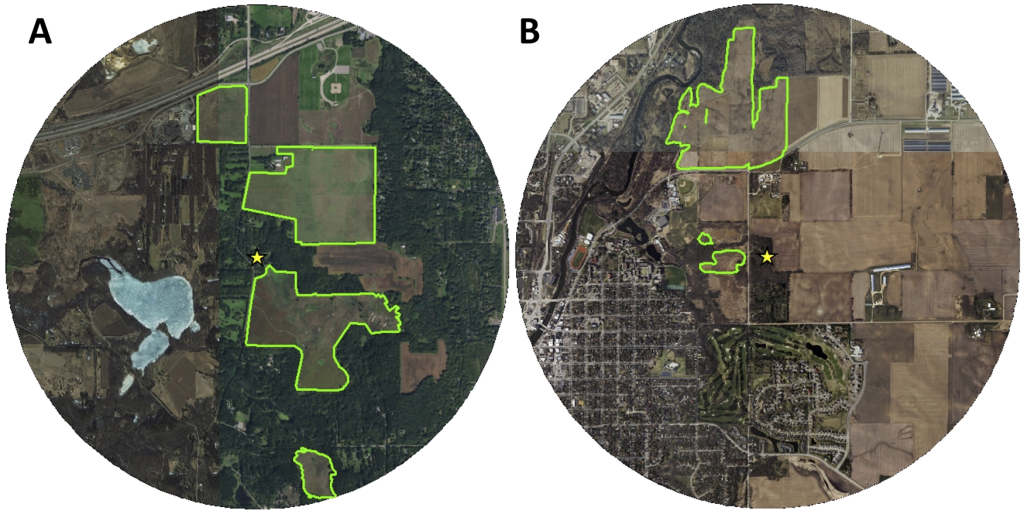
A) Консерватория Белвин. Б) Дендрарий Каулинга при Карлтонском колледже. Зеленые контуры показывают края восстановленных прерий на каждом участке. Мы немного изменили края прерий в Cowling Arboretum (Карлтонский колледж) на основе шейпфайлов посадочных единиц 2010 года, созданных Вей-Хсином Фу и Нэнси Бракер с помощью ArcGIS версии 10.0. Желтые звезды обозначают места расположения ульев. Карты простираются на 2 км вокруг ульев. Источники базового слоя карты World Imagery: Esri, DigitalGlobe, GeoEye, i-cubed, USDA FSA, USGS, AEX, Getmapping, Aerogrid, IGN, IGP, swisstopo, и сообщество пользователей ГИС.
2.3 Флористические исследования
Раз в две недели в период наблюдения за колониями мы проводили цветочные исследования на каждом из прилегающих участков реконструированной прерии вокруг колоний. Обследования заключались в выявлении всех цветущих соцветий в квадратных метрах. Мы размещали квадраты на десяти случайно выбранных расстояниях (функция randbetween, Excel) вдоль 200-метровых трансектов, проходящих через каждый участок прерии. Когда мы замечали, что определенные виды цветущих цветов в изобилии присутствуют в прериях, но отсутствуют в наших квадратах, мы регистрировали эти виды как цветущие в эту дату отбора проб. Мы определяли цветущие травы как можно точнее, почти во всех случаях с точностью до вида.
2.4 Управление колониями медоносных пчел
2.4.1 Наблюдательные ульи и сараи
Деревянные навесы защищали ульи от ветра и прямых солнечных лучей. Ульи освещались люминесцентными или светодиодными лампами. Мы размещали колонии в наблюдательных ульях Ольстерского типа (Mann Lake, https://www.mannlakeltd.com/observation-hive) с коробкой внизу, вмещающей 5 рамок пчелиного воска, и одной видимой рамкой между двумя стеклами сверху. Мы модифицировали ульи для наблюдения, чтобы уменьшить пространство вокруг видимой рамки примерно до одного пчелиного места (~1 см, [58]), чтобы предотвратить образование рабочими нескольких слоев и заслонение друг друга от глаз. Мы запечатали нижний вход в улей и просверлили отверстие диаметром 2,54 см в дереве на полпути вверх по видимой рамке, чтобы можно было вставить пластиковую трубку через стенку наблюдательного улья в улей, позволяя фуражирам входить и выходить только на видимую рамку. Учитывая, что фуражиры предпочитают танцевать недалеко от входа в улей [26,59], мы добавили барьеры, чтобы фуражирам нужно было пройти дальше в улей, чтобы добраться до мест, не видимых камерами, обеспечивая запись танцевальной площадки. Чтобы предотвратить спуск возвращающихся фуражиров с видимых рамок на рамки ниже в пределах первых 34 см от входа, мы использовали стальные скобяные изделия (расстояние между ними 0,32 см). Чтобы предотвратить перемещение фуражиров на противоположную сторону видимой рамки в пределах 34 см от входа, мы использовали металлические барьеры (Mann Lake, https://www.mannlakeltd.com/metal-frame-rests-10-pack).
2.4.2 Управление колониями
В течение каждого года наблюдений мы размещали по три колонии на каждом участке. Все колонии мы создавали из 2-фунтовых пакетов рабочих с молодыми спаренными «итальянскими» матерями (полученными в основном от европейского подвида Apis mellifera ligustica), приобретенных у коммерческого пчеловода в Северной Калифорнии (https://www.ohbees.com/). Чтобы убедиться, что все колонии имеют схожие по размеру кормовые силы, запасы пищи и потребности в выращивании расплода, мы наполнили каждый улей для наблюдения королевой, приблизительно 6 000-8 000 рабочих, двумя рамками с пчелиным воском, заполненными закрытым расплодом (куколками), одной рамкой с незакрытым расплодом (личинками), одной рамкой с медом/пергой и двумя вытянутыми пустыми рамками с воском.
Раз в неделю мы осматривали колонии на наличие признаков болезни, проблем с королевой и/или скученности. Если колонии казались слишком тесными, мы удаляли рамки с расплодом. Когда королеве требовалось больше места для откладки яиц, мы удаляли рамки с кормом. Когда мы удаляли эти рамки, мы заменяли их пустыми, затянутыми гребнями. Поскольку оказалось, что фуражиры предпочитают танцевать на рамках с постоянным прикрытием других рабочих, всякий раз, когда большинство куколок выходили взрослыми из верхней, видимой рамки, мы меняли эту рамку на рамку с расплодом снизу, поощряя больше рабочих проводить время на видимой рамке. В двух случаях, когда осмотр показал, что королева умерла или перестала откладывать яйца (оба случая произошли в 2017 году на участке Cowling Arboretum), мы заменили королеву новой, спаренной «итальянской» королевой. Мы проводили инспекции в конце последнего наблюдения в течение каждой недели, чтобы дать колониям время адаптироваться к любым изменениям до начала следующей недели наблюдений. Когда видеозапись не велась, мы держали стеклянные стенки наблюдательных ульев покрытыми изоляционными панелями из пенопласта, чтобы помочь колониям терморегуляции.
2.5 Видеосъемка
Мы проводили видеосъемку танцплощадки (участок воскового гребня возле входа, где происходит большинство танцев) один день в неделю в Belwin Conservancy и два дня в неделю в Cowling Arboretum с 50-минутными записями на колонию в день. Дни чередовались между утренними (8:00-12:00) и дневными (13:00-17:00) записями, и мы записывали колонии в случайном порядке каждый день. В заповеднике Белвин все записи производились с помощью камеры SONY HDR-CX440. В дендропарке Каулинг все записи производились камерой SONY FDR-AX33. В обоих случаях мы разместили камеры на расстоянии примерно 75 см от танцплощадки и настроили их на фокусировку на участке шириной примерно 30 см х 25 см, прилегающем к входу в колонию. Во время записи наблюдатель указывал на всех танцующих и отмечал, есть ли у них видимый пыльцевой налет.
2.6 Извлечение пространственной информации из танца виляния
2.6.1 Выбор танцев виляния для анализа
Чтобы отобрать примерно равное количество танцев из всех колоний и за все недели кормового сезона, мы расшифровали первые 10 танцев на нектар и первые 10 танцев на пыльцу. Танцоров с пыльцой мы отнесли к танцорам пыльцы, а танцоров без пыльцы — к танцорам нектара. Возможно, что некоторые из этих танцев могли быть рекламой источников воды или смолы. Однако в обычных условиях лишь незначительное меньшинство фуражиров собирает воду [26,55,60] или смолы [61,62,63]. Таким образом, вполне вероятно, что большинство танцоров без груза на задних ногах рекламировали источники нектара, а большинство танцоров с грузом на задних ногах — источники пыльцы. Стоит отметить, что танцоры, рекламирующие пыльцу, могли также собирать нектар во время той же поездки, возможно, с тех же видов растений. Большинство видеороликов (в среднем 57,5%) содержали не менее 10 танцев, рекламирующих источники нектара. Менее половины видеороликов содержали 10 танцев, рекламирующих источники пыльцы (в среднем 22,5%). В видео с менее чем 10 танцами, рекламирующими один из двух пищевых ресурсов, мы декодировали все танцы для этого ресурса, в среднем 8,4±0,4 танцев для нектара и 4,7±0,3 танцев для пыльцы на видео в Belwin Conservancy и 8,3±0,5 танцев для нектара и 6,8±0,5 танцев для пыльцы на видео в Carleton College.
2.6.2 Расшифровка танцев виляния
Для картирования танцев виляния необходимо сначала измерить средний угол и продолжительность виляния в каждом танце. Мы измерили эти две переменные в соответствии с методами Кувиллона и др. [64], за исключением того, что для расчета продолжительности каждого виляния использовался Quicktime 7, а не FinalCutExpress. Как только мы наблюдали танец, мы выбирали серию из как минимум 6 последовательных виляний и избегали декодирования первого и последнего виляний в этой серии. Это позволило нам рассчитать средний угол и продолжительность четырех последовательных виляний, что, как было показано, тесно коррелирует со средним углом и продолжительностью всех виляний в танце [64]. Так как танцоры виляния регулируют углы наклона относительно силы тяжести, углы измерялись относительно вертикальных отвесов (белая обивочная нить, прикрепленная к вольфрамовым грузикам, подвешенным к гвоздям в дереве в верхней части видимой рамы на расстоянии 10 см друг от друга). Азимут солнца во время каждого танца рассчитывался с помощью подпрограммы R SunPosition (согласно Schürch и др. [56]) с дополнительным смещением на 5 часов от среднего времени по Гринвичу (центральное стандартное время во время летнего перехода).
2.6.3 Моделирование облаков плотности вероятности на основе информации, декодированной из танцев
Поскольку вариации между танцующими пчелами и между виляниями внутри танца делают точное местоположение рекламируемого цветочного участка неопределенным, мы отобразили декодированные танцы, используя оценки этой неопределенности в соответствии с методами [56]. Для каждого декодированного танца мы использовали байесовскую линейную калибровочную модель для создания 1000 расстояний, сосредоточенных вокруг среднего расстояния, на которое указывает продолжительность танца. Мы определили дисперсию, используя набор калибровочных данных, полученных при обучении фуражиров медоносных пчел посещать кормушки с сахарной водой на разных расстояниях и расшифровывать их танцы. Поскольку калибровочные данные из [56] не показали значительного отличия связи между продолжительностью виляния и расстоянием от аналогичных данных, собранных нами в заповеднике Белвин в сентябре 2015 года (перехват: t(122) = 0,825, p = 0,410; наклон: t(122) = 0,618, p = 0,537), мы объединили оба набора калибровочных данных для получения расчетных расстояний. Объединенный набор калибровочных данных показывает взаимосвязь между продолжительностью танца в секундах (d) и расстоянием в метрах (D):
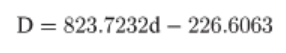
Кроме того, мы использовали средний угловой разброс калибровочных данных из Belwin Conservancy (распределение фон Мизеса, каппа = 35,4 на основе разброса танцев для самого дальнего фидера в Belwin Conservancy) для создания 1000 заголовков, центрированных вокруг среднего заголовка танца. Объединив расчетные расстояния и направления, мы нанесли на карту 1000 точек для каждого танца, что позволило нам рассчитать долю точек, которые попали внутрь прерий, и, таким образом, вероятность того, что каждый танец рекламировал цветочные пятна в реконструированных прериях.
2.6.4 Анализ количества перебежек в танце
Танцы с вилянием часто включают несколько схваток, каждая из которых включает некоторое количество последовательных циклов (фазы виляния и возвращения), разделенных периодами обмена пищей или хождения в другую область танцевальной площадки. Поэтому, чтобы определить оценку фуражирами прибыльности прерийных и не прерийных источников нектара, мы отследили нектарных танцоров, записанных в Belwin Conservancy, до первого кадра, на котором танцор вошел в улей, а затем вперед до последнего кадра, на котором танцор был виден. В 352 случаях из 401 мы могли отследить нектарную танцовщицу с того момента, как она вошла в зону танцпола, до того момента, как она покинула его. Мы подсчитали общее количество виляний во всех схватках этих танцев.
2.7 Сбор и идентификация пыльцы
Чтобы оценить, рекламируют ли медоносные пчелы пыльцу местных прерий, и определить, какие таксоны они рекламируют, мы взяли пыльцевые грузы у подгруппы танцующих фуражиров. В один из дней видеосъемки каждую неделю мы заменяли стекло улья на кусок оргстекла, в котором просверливали четыре круглых отверстия диаметром 8-10 см. На каждое отверстие был наклеен кусок прозрачного ацетатного листа, чтобы можно было поднимать лист как дверь. Чтобы собрать данные о наибольшем количестве различных цветочных пятен, когда мы наблюдали танцовщицу с пыльцой, мы классифицировали ее танец по углу (с точностью до 30 градусов) и цвету пыльцы (разные цвета пыльцы обычно указывают на разные виды цветов, [65]). Фуражиры медоносных пчел редко собирают пыльцу более чем с одного вида цветущих растений во время одной фуражировки [66]. Каждый раз, когда возникал танец для новой комбинации угла/цвета, мы рассматривали его как уникальный участок цветов. Для каждого уникального участка мы позволили одному танцору завершить 6 кругов, а затем поймали его с помощью трубчатой сетчатой клетки, которую мы поместили вокруг него, пока он не вошел в нее. Затем мы закрыли клетку крышкой и поместили ее на лед. Как только танцовщица теряла сознание под воздействием низких температур (5-10 минут), мы ненадолго вынимали ее из клетки и собирали пыльцу для последующего анализа. После завершения 50-минутного периода регистрации мы помещали фуражиров в клетку снаружи и позволяли им улететь обратно в свои колонии.
Чтобы облегчить идентификацию пыльцы путем удаления содержимого пыльцы, липидов и других остатков, мы ацетолизировали все пыльцевые грузы — процесс, включающий воздействие нагретой смеси кислот [49]. Затем мы рассмотрели оставшиеся спорополлениновые стенки пыльцевых зерен с помощью светового микроскопа при 400-кратном увеличении и сравнили их с эталонными образцами пыльцы известных видов. Каждый раз, когда мы встречали новые виды/роды во время флористических исследований в Belwin Conservancy, мы собирали пыльцу и добавляли ее в нашу библиотеку пыльцевых эталонов. По возможности мы распределяли пыльцу таксонов по следующим категориям: пыльца таксонов, являющихся родными для экосистем прерий и присутствующих в реконструированных прериях (таблица S1, рисунок S1, [67]), пыльца таксонов, являющихся родными для региона Верхнего Среднего Запада, но не для экосистем прерий (например, деревья, лианы, лесные травы), и пыльца таксонов, не являющихся родными для Верхнего Среднего Запада (на основе базы данных USDA PLANTS plants.sc.egov.usda.gov). Мы расшифровали танцы и нанесли на карту источники этих типов пыльцы.
Из 335 собранных пыльцевых масс мы разрезали 104 пополам, чтобы рассмотреть одну половину под микроскопом и секвенировать область ITS (рибосомальный внутренний транскрибируемый спейсер) другой половины (праймеры: ITS4a и ITS5, [68]) для подтверждения микроскопических идентификаций и обеспечения более конкретной таксономической идентификации в некоторых случаях. В случаях, когда результаты ITS BLAST указывали на грибковые таксоны, мы также секвенировали хлоропластный ген, rbcL (большая субъединица RuBisCO; праймеры были в соответствии с [69]). Мы выделяли ДНК с помощью 5 мм боросиликатных шариков, Qiagen TissueLyser II и Qiagen DNeasy Plant Mini-Kits. Мы амплифицировали ДНК-маркеры с помощью GE Healthcare Life Sciences illustra PuReTaq Ready-To-Go PCR Beads с 1 мкл ДНК, 2,5 мл 10нМ каждого праймера и 19 мкл воды, с настройками термоциклера согласно [68] для области ITS и [69] для rbcL. Секвенирование по методу Сэнгера проводили на анализаторе ДНК Applied Biosystems 3730xl. Мы очистили полученные последовательности в программе Geneious и провели BLAST по базе данных NCBI. Мы отнесли пыльцу к таксонам, основываясь на самом низком таксономическом уровне, который разделяли 10 лучших BLAST-попаданий для ITS (или, в 10 случаях, rbcL). Результаты секвенирования, полученные в данном исследовании, мы поместили в базу данных GenBank под номерами доступа MN273343-MN273438 (последовательности ITS) и MN284913-MN284922 (последовательности rbcL).
2.8 Статистический анализ
2.8.1 Доля танцевальных рекламных площадок в прериях в течение кормового сезона
Мы провели все статистические тесты с использованием статистического программного обеспечения R (версия 3.6.0). После нанесения 1000 точек на танцы мы рассчитали 95% доверительный интервал Агрести-Кулла [70,71] для доли танцев в прериях, где p — число танцев в прериях, а n — общее число танцев (в обоих случаях масштабированное до фактического числа танцев путем деления числа смоделированных точек на 1000). Кроме того, мы использовали хи-квадрат тест Пирсона (chisq.test), чтобы проверить, различаются ли доли танцев для участков прерий в разные части кормового сезона (май/июнь, июль и август/сентябрь) на каждом участке и для каждого типа пищи (пыльца и нектар) с поправкой Бонферрони для множественных тестов. Когда тесты хи-квадрат показывали значительное влияние сезона, мы использовали смешанную модель логистических регрессий с сезоном в качестве фиксированного эффекта и колонией в качестве случайного эффекта, взвешенного по количеству танцев (glmer(Proportions_in_prairies ~ Season + (1|Colony), family = binomial(link = «logit»), weights = Dances), чтобы получить p-значения для сравнения между частями сезона.
2.8.2 Взаимосвязь между вероятностью того, что нектарный танец рекламировал источники в прериях, сезоном и количеством перелетов на танец
Для танцев, рекламирующих источники нектара в Belwin Conservancy, мы оценили взаимосвязь между количеством виляний на танец, частью сезона и вероятностью того, что танец рекламирует цветочный участок в прерии, используя линейную модель (lm). Поскольку не было обнаружено значительного влияния того, рекламировал ли танец участок прерии, на количество покачиваний, которые выполнял танцор, мы далее проанализировали влияние только сезона, используя ANOVA (aov) и тест на достоверность значимых различий Тьюки (TukeyHSD).
2.8.3 Сезонные изменения среднего расстояния до рекламируемых источников нектара
Мы рассчитали расстояния до рекламируемых источников нектара, используя среднее соотношение между продолжительностью и расстоянием из калибровочных данных (см. уравнение в разделе 2.5.3), и сравнили эти расстояния с помощью ANOVA (aov). Мы оценили различия в среднем расстоянии до рекламируемых источников в разные периоды кормового сезона с помощью теста Тьюки HSD (TukeyHSD).
3. Результаты
3.1 Танцы, рекламирующие места в прериях в течение всего кормового сезона
В заповеднике Белвин до августа/сентября расчетная доля танцев, рекламирующих источники нектара в прериях, была ниже 20%. В августе и сентябре 32,2% танцев, рекламирующих источники нектара, относились к местам в реконструированных прериях (рис. 3 и 2), что свидетельствует о статистически значимом сезонном изменении (рис. 3; табл. 1). Анализ логистической регрессии смешанной модели показал значительные различия между маем/июнем и августом/сентябрем (z = -3,57, p<0,001) и между июлем и августом/сентябрем (z = -2,48, p = 0,013), но не между маем/июнем и июлем (z = -0,91, p = 0,36). Аналогичная сезонная тенденция наблюдалась для доли танцев, рекламирующих источники пыльцы в Belwin Conservancy: 15,9% танцев рекламировали места в прериях в мае/июне, 12,0% в июле и 24,8% в августе/сентябре, хотя она не была значительной (Рис. 3; Таблица 1). Напротив, доля танцующих, рекламирующих источники нектара и пыльцы в прериях в Карлтонском колледже, составляла менее 10% в течение всего кормового сезона и существенно не менялась от месяца к месяцу (рис. 3; табл. 1). Ошибка в информации о расстоянии и направлении в танцах могла повлиять на наши оценки доли танцев, которые рекламировали места в прериях, несколько по-разному в двух местах. Возможно, что наши оценки в Карлтонском колледже были ниже, чем в Belwin Conservancy, отчасти из-за меньшего размера ближайших участков прерий в Карлтонском колледже. Несмотря на это потенциальное смещение, размер нашей выборки в 904 танца должен был быть достаточным для выявления сезонной тенденции в Карлтонском колледже.
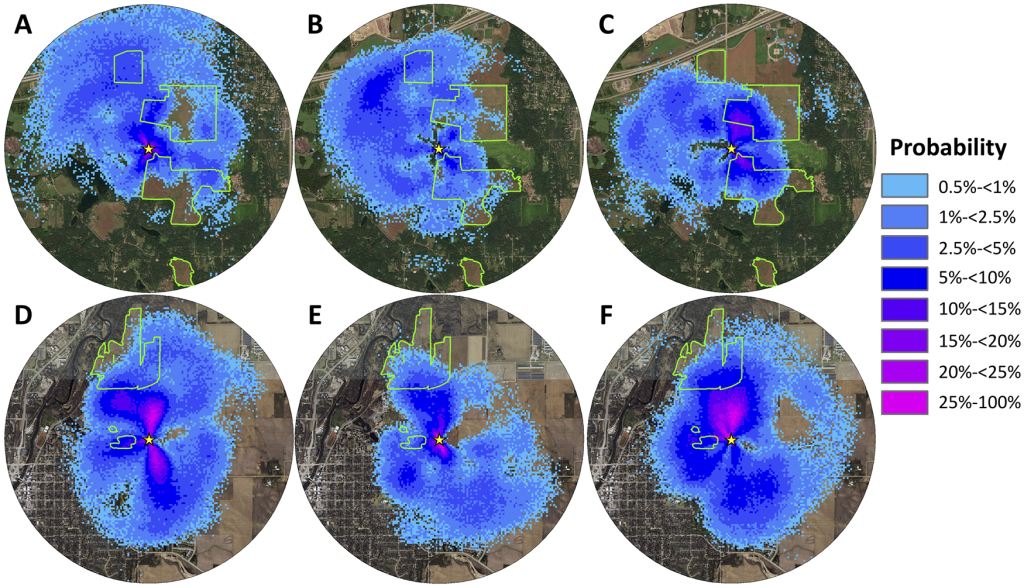
Цвета показывают вероятность того, что хотя бы один танец рекламировал цветочный участок в пределах квадрата сетки 5 м х 5 м. Зеленые контуры показывают края восстановленных прерий на каждом участке. Желтые звезды обозначают места расположения ульев. Карты простираются на 2 км вокруг ульев.
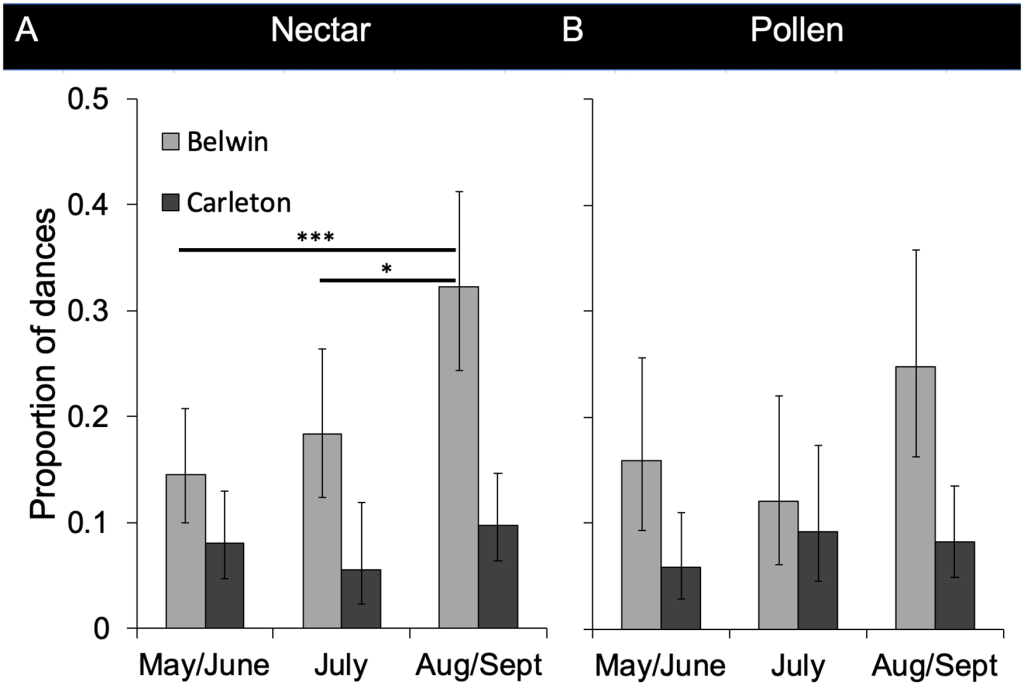
A) Источники нектара в Belwin Conservancy (май/июнь: n = 166 танцев, июль: n = 119, август/сентябрь: n = 116) и Carleton College (май/июнь: n = 180 танцев, июль: n = 109, август/сент: n = 207). B) Источники пыльцы в Belwin Conservancy (май/июнь: n = 80 танцев, июль: n = 69, август/септ: n = 74) и Carleton College (май/июнь: n = 149 танцев, июль: n = 89, август/септ: n = 170). Пропорции основаны на расположении смоделированных точек. Планки ошибок показывают 95% доверительные интервалы Агрести-Коула, где p — количество танцев в прериях, а n — общее количество танцев, увеличенное до фактического количества танцев (количество смоделированных точек, деленное на 1000). * указывает на p<0,05, *** указывает на p<0,001. Источники базового слоя карты World Imagery: Esri, DigitalGlobe, GeoEye, i-cubed, USDA FSA, USGS, AEX, Getmapping, Aerogrid, IGN, IGP, swisstopo, и сообщество пользователей ГИС.
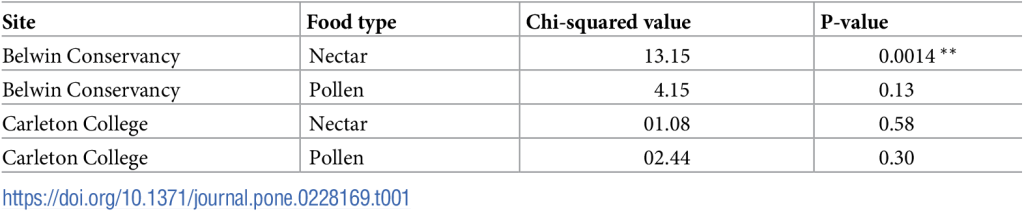
Чтобы лучше понять сезонные изменения в Belwin Conservancy, мы посмотрели, есть ли различия между танцами, которые, вероятно, рекламировали места прерий и танцами, которые, вероятно, не рекламировали места прерий (доля смоделированных точек, которые попали в прерии). В частности, мы смотрели на количество раз, которое танцор повторял информацию о местоположении во время танца (общее количество выполненных виляний). Мы не обнаружили значительного влияния вероятности того, что данный нектарный танец рекламировал местоположение прерии, на общее количество виляний в этом танце (df = 346, t = 0,734, p = 0,46), а также взаимодействия между сезоном и вероятностью того, что танец рекламировал местоположение прерии (p = 0,30 и p = 0,46). Однако мы увидели значительное влияние сезона на длину танца как в модели с взаимодействиями (t = 2,86, p<0,01), так и в последующем ANOVA с фиксированным эффектом только сезона (F(2,349) = 4,84, p<0,01; рис. 4). Танцы в июле (33,43±3,22 виляний, среднее±SEM) включали значительно большее количество виляний, чем танцы в мае/июне (25,92±3,00 виляний; p<0,05) или августе/сентябре (24,23±2,32 виляний; p<0,05).
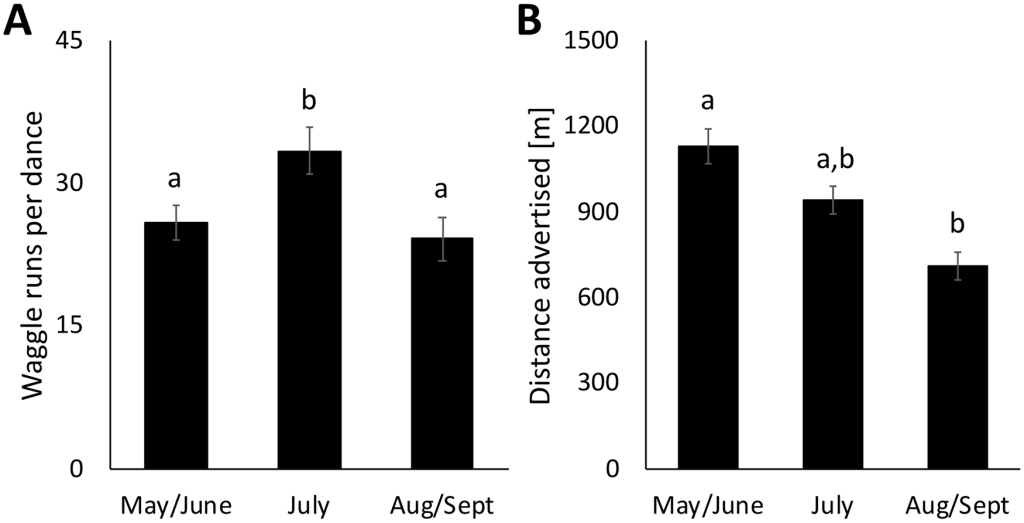
Столбики ошибок показывают стандартную ошибку среднего значения. Буквы указывают на значительные различия (p<0,05). Размеры выборки: пробеги — май/июнь (n = 147), июль (n = 106) и август/сентябрь (n = 99); расстояния — май/июнь (n = 166), июль (n = 119) и август/сентябрь (n = 116).
Мы также сравнили средние расстояния до рекламируемых источников нектара в течение кормового сезона. В заповеднике Белвин танцоры рекламировали источники нектара, которые находились в среднем на расстоянии 954,27±33,73 м (среднее±SEM) от улья, а максимальное рекламируемое расстояние составило 4 873,61 м. Однако в течение сезона наблюдалось значительное снижение расстояния от 1 129,72±62,24 в мае и июне до 713,30±49,62 в августе и сентябре (F(2,398) = 13,83, p<0,0001; рис. 4). В Карлтонском колледже среднее расстояние до рекламируемых источников нектара составило 817,57±34,73 и существенно не изменилось в течение сезона (F(2,381) = 0,45, p = 0,64).
Средние значения и тенденции для танцев, рекламирующих источники пыльцы, были очень похожи на те, что наблюдались для источников нектара. Пыльцевые танцы в заповеднике Белвин рекламировали источники на среднем расстоянии 927,35±40,84 метров со значительным сезонным снижением с мая по август (F(2,220) = 5,79, p<0,01). Пыльцевые пляски в Карлтонском колледже рекламировались на среднем расстоянии 830,91±32,56 метров без значительного сезонного изменения (F(2,329) = 0,80, p = 0,45).
3.2 Таксоны рекламируемых источников пыльцы
В этом исследовании было зафиксировано, что фуражиры танцевали, привлекая птенцов для сбора пыльцы по меньшей мере 34 различных родов из 25 семейств, включая семь местных таксонов прерий (рис. 5; таблица S1). Танцоры рекламировали эти местные таксоны прерий в июле, августе и сентябре (Таблица S1). Из 335 собранных пыльцевых масс 9,2% принадлежали таксонам коренных прерий, 9,9% — таксонам коренных прерий, но из не прерийных экосистем (леса, водно-болотные угодья и т.д.), 58,8% — не коренным таксонам, а 22,1% не могли быть идентифицированы достаточно точно, чтобы определить, являются ли они коренными для региона или нет (Рис. 5; Таблица S1). При исследовании цветов был зафиксирован 21 вид местных цветов прерий, цветущих в Belwin Conservancy и 10 в Carleton College (S1 рис.). Они включали пять из семи рекламируемых таксонов местных прерий, включая Solidago spp., Agastache spp., Dalea candida, Dalea purpurea и Rudbeckia spp.
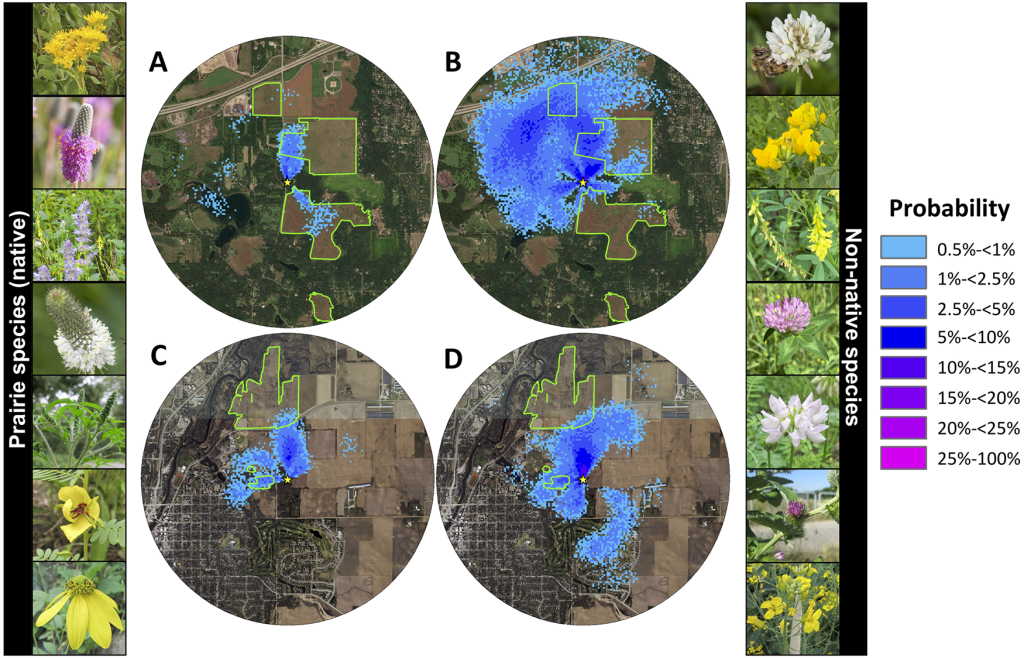
Цвета показывают вероятность того, что хотя бы один танец рекламировал цветочный участок в пределах квадрата сетки 5 м х 5 м. Зеленые контуры показывают края восстановленных прерий на каждом участке. Желтые звезды обозначают места расположения ульев. Карты простираются на 2 км вокруг ульев. На фотографиях слева показаны все подтвержденные рекламой таксоны местных прерий: Solidago sp., Dalea purpurea, Agastache sp., Dalea candida, Ambrosia sp., Chamaecrista fasciculata, и представитель трибы Heliantheae (вероятно, Rudbeckia sp.). На фотографиях справа показаны семь наиболее рекламируемых таксонов неместных цветов: Trifolium repens/hybridum, Lotus corniculatus, Melilotus officinalis, Trifolium pretense/incarnatum, Securigera varia*, Arctium minus и Brassica sp. *Кредиты изображений: Хизер Холм. Источники базового слоя карты World Imagery: Esri, DigitalGlobe, GeoEye, i-cubed, USDA FSA, USGS, AEX, Getmapping, Aerogrid, IGN, IGP, swisstopo, и сообщество пользователей ГИС.
4. Обсуждение
Результаты нашего исследования дали ответы на два основных вопроса о том, как ведут себя фуражиры медоносных пчел, когда им предоставляется доступ к большим реконструированным прериям. Во-первых, хотя большинство танцев рекламировали источники пищи, не связанные с прериями, в консерватории Белвин мы наблюдали значительное увеличение доли танцев, посвященных источникам нектара в прериях в конце кормового сезона. Во-вторых, мы установили, что медоносные пчелы воспринимают семь таксонов местных цветов прерий как выгодные источники пыльцы и рекламируют их с помощью танцев: Solidago spp., Dalea purpurea, Agastache sp., Dalea candida, Ambrosia spp., Chamaecrista fasciculata и представитель трибы Heliantheae (вероятно, вид Rudbeckia, судя по наибольшему количеству BLAST-попаданий).
Чтобы лучше понять сезонные изменения в доле нектарных танцев, рекламирующих участки прерий в Belwin Conservancy, мы определили, совершают ли фуражиры, рекламирующие источники нектара в прериях, больше или меньше виляющих пробежек, чем фуражиры, рекламирующие источники нектара вне прерий в течение фуражировочного сезона. Мы не обнаружили значительной связи между вероятностью того, что танец рекламировал место в прериях, и количеством выполненных виляний. Значительно большее число покачиваний в поисках источников нектара в прериях в августе/сентябре поддержало бы идею о том, что фуражиры воспринимают цветы прерий в конце сезона как более выгодные источники нектара, чем цветы прерий в конце сезона. Отсутствие различий предполагает, что увеличение доли танцев в августе/сентябре может отражать изменения в изобилии полезных цветов внутри прерий по сравнению с другими районами. Тем не менее, мы заметили значительное влияние сезона: в июле были самые продолжительные танцы. Большее число виляний указывает на то, что в этот период фуражиры тратили больше энергии на привлечение других фуражиров, что может быть результатом того, что фуражиры воспринимали июльские источники нектара как более выгодные или воспринимали большую потребность колонии в нектаре или способность запасать его в июле [72]. У новобранцев, которые следуют за большим количеством виляний, значительно больше шансов найти источник пищи [73], поэтому увеличение в среднем на 7-9 виляний в июле потенциально может привести к увеличению успеха рекрутирования в среднем на один танец. Поскольку каждый новобранец, скорее всего, также будет танцевать, эта разница в количестве виляний в расчете на один танец, вероятно, увеличит рекрутирование на этом участке экспоненциально, поскольку больше фуражиров будут его рекламировать [74].
Мы также проанализировали средние расстояния до рекламируемых источников нектара в течение сезона в Belwin Conservancy и обнаружили значительные сезонные изменения: меньшие расстояния рекламируются в более поздний сезон. Средние расстояния, которые мы обнаружили, находятся в пределах диапазона предыдущих исследований [36,43,71,75]. Однако уменьшение рекламируемых расстояний удивительно, поскольку многочисленные исследования показали, что колонии медоносных пчел на верхнем Среднем Западе и в соседних штатах сталкиваются с периодом обеднения, когда в конце лета и осенью наблюдается меньшее разнообразие цветущих цветов (Северная Дакота: [76]; Огайо: [75]; Мичиган: [77]; Айова: [78]). Период с начала июля до начала августа обычно является временем года, когда колонии медоносных пчел в Миннесоте набирают наибольший вес и производят наибольшее количество меда [79,80]. Период нектарового голодания обычно приводит к увеличению расстояния, на которое ведется реклама [27,40,41,42]. Наши результаты показывают, что нахождение вблизи реконструированной прерии в поздний сезон может дать колониям значительное преимущество в плане приспособленности, позволяя им добывать корм ближе к дому и тратить меньше энергии на одну поездку в то время года, когда в противном случае им пришлось бы путешествовать гораздо дальше. Недавнее исследование показало, что колонии в ландшафтах с преобладанием кукурузы и сои теряют вес, а их пчелы-воспитательницы теряют запасы жира, начиная с августа, но перемещение колоний в большую прерию в августе избавило их от этих последствий [78]. Более низкая дальность полета, которую мы наблюдали в августе/сентябре, потенциально может способствовать этому эффекту спасения.
Существует несколько возможных объяснений того, почему фуражиры в наших колониях показали сезонные изменения в доле танцев виляния в поисках источников нектара в прериях в Belwin Conservancy, но не в Carleton College. Различия в погоде между этими двумя участками могли по-разному повлиять на производство нектара местными видами прерий и видами, не относящимися к прериям. Температура и количество осадков могут оказывать значительное влияние на время цветения и нектаропродуктивность многих видов травянистых растений [81,82,83]. Среднее расстояние между нашими колониями и реконструированными прериями в Belwin Conservancy было меньше, чем среднее расстояние между нашими колониями и реконструированными прериями в Carleton College (рис. 1), что делает их более легкими для перелета и, следовательно, более привлекательными [71]. Мы также отметили большее разнообразие цветущих растений в Belwin Conservancy, включая Solidago rigida, цветущую в более поздний сезон, которые могли быть особенно привлекательными источниками нектара и пыльцы в это время года (S1 рис.). К сожалению, наши методы идентификации пыльцы не позволяют различать виды рода Solidago, но они выявили этот род как один из наиболее часто рекламируемых источников пыльцы местных прерий (Таблица S1). Виды Solidago имеют тенденцию расти большими, плотными скоплениями из-за их способности производить колонии клонов с помощью корневищ [84]. Такой характер роста может сделать участки солидаго особенно привлекательными для медоносных пчел, поскольку на один и тот же участок цветов можно привлечь много кормильцев, не истощая его кормом. Возможно, что растения солидаго вокруг Belwin Conservancy были более сконцентрированы в восстановленных прериях, в то время как большая часть растений солидаго, цветущих рядом с нашими колониями в Карлтонском колледже, росла на обочинах дорог или в других местах обитания, не связанных с прериями. Учитывая, что мы могли исследовать цветы только в прериях, мы не можем быть уверены в этом.
В дополнение к семи таксонам прерий, фуражиры в нашем исследовании рекламировали разнообразный набор неместных и местных пыльц из не прерийных местообитаний. Тремя наиболее часто рекламируемыми таксонами пыльцы были Trifolium repens/hybridum, Melilotus officinalis и Lotus corniculatus (Таблица S1). Эти таксоны являются неродными травами из Европы, завезенными в Северную Америку в качестве кормовых культур для скота, и с тех пор они стали обычными сорняками [85]. Все три вида имеют длительный период цветения, приходящийся на июль (S1 рис.), что может помочь объяснить большое количество рекламируемых пятен. Тот факт, что медоносные пчелы также были завезены в Северную Америку из Европы, поднимает вопрос о том, могли ли медоносные пчелы выработать сильные предпочтения к сигналам от цветов в их родном ареале. С другой стороны, успех европейских медоносных пчел на шести континентах и тот факт, что медоносные пчелы являются наиболее часто регистрируемыми цветочными посетителями в естественной среде обитания по всему миру [86], предполагает, что у них очень пластичные кормовые предпочтения. Танцы рекламировали ряд других неместных сорных таксонов, включая Trifolium pratense/incarnatum, Securigera varia, Arctium minus и Brassica spp. (Таблица S1). Эти неместные таксоны совпадают с таксонами, найденными в пыльце, собранной колониями медоносных пчел в отдельном близлежащем исследовании, за исключением Arctium minus и Lotus corniculatus [13,76]. Карты танцев, рекламирующих их, показывают, что большинство, но не все, танцы для этих таксонов были для участков за пределами прерий (рис. 5). Обследования цветов подтвердили, что ряд неместных видов цвели в прериях на обоих участках (S1 рис.). Кроме того, танцы в мае, июне и июле рекламировали ряд местных таксонов, не относящихся к прериям, включая древесные виды таких родов, как Gleditsia, Rhus, Cornus, Tilia и Parthenocissus, которые обеспечивают кормушки медоносных пчел очень концентрированными пятнами цветов (S1 Таблица).
5.Заключение
Хотя мы не нашли доказательств того, что реконструированные прерии представляют собой очень привлекательный ресурс для медоносных пчел в мае, июне и июле, мы нашли доказательства того, что реконструированные прерии могут стать очень привлекательными в более поздний сезон, что потенциально может привести к значительным преимуществам для здоровья колоний медоносных пчел. Кроме того, мы обнаружили, что пчелы-фуражиры воспринимают семь местных прерийных таксонов, найденных в реконструированных прериях, как достойные рекламы для своих сородичей. Наши результаты показывают, что включение этих таксонов, особенно Dalea purpurea, Dalea candida и Agastache sp. в высокой плотности может сделать прерии более привлекательными для медоносных пчел в июле, что является полезной информацией как для землеустроителей, которые хотят обеспечить корм для медоносных пчел, так и для тех, кто хочет избежать потенциальной конкуренции между медоносными и местными пчелами на своей земле. В случаях, когда землевладельцы обеспокоены конкуренцией [87], они могут либо избегать посадки этих видов с высокой плотностью, либо, если их целью является сохранение специализированных пчел, которые полагаются на эти виды для получения пыльцы (например, специалисты Colletes на Dalea [88]), они могут ограничить количество колоний медоносных пчел, имеющих доступ к посадкам в прерии. Кроме того, наши результаты показывают, что в настоящее время несколько видов неместных источников пыльцы, включая виды из рода Trifolium, Melilotus officinalis и Lotus corniculatus, имеют большое значение для колоний медоносных пчел. В настоящее время мы изучаем полный рацион питания колоний медоносных пчел, расположенных вблизи восстановленных прерий, чтобы получить больше информации о наиболее привлекательных видах прерий для дружественных медоносным пчелам посадок. Будущие эксперименты, включающие посадку тщательно контролируемых участков цветов и наблюдение за поведением медоносных пчел, помогут определить, насколько важны площадь участка и плотность посадки для привлечения медоносных пчел-фуражиров.
Вспомогательная информация
Данные S1. Продолжительность танцев для кормушек с сиропом на разных расстояниях от наблюдательных ульев в Belwin Conservancy в 2015 году.
(CSV)
S2 Данные. Файл значений, разделенных запятыми, с данными 1528 танцев виляния, расшифрованных в консерватории Белвин и Карлтонском колледже.
(CSV)
S3 Данные. Файл с разделенными запятыми значениями общего числа виляний в танцах в поисках источников нектара в заповеднике Белвин.
(CSV)
S4 Данные. Файл значений, разделенных запятыми, с оценкой расстояний до источников нектара и пыльцы, рекламируемых фуражирами в заповеднике Белвин.
(CSV)
S5 Данные. Файл значений, разделенных запятыми, с данными идентификации пыльцы и расшифровки танцев с обоих участков.
(CSV)
S6 Данные. Файл значений, разделенных запятыми, с указанием участков, дат и доли исследованных квадратов, в которых мы наблюдали каждый вид форбса.
(CSV)
S1 Таблица. Пыльцевые массы, собранные с танцующих вихляющих фуражиров.
Пыльца была собрана с каждого танца, рекламирующего уникальную комбинацию цвета пыльцы и угла танца, чтобы определить таксон каждого цветочного пятна, рекламируемого в течение 50-минутного еженедельного периода отбора проб. Родные таксоны прерий отмечены символом звездочки (*), а неродные таксоны отмечены символом плюса (+).
(DOCX)
S1 Рис. Диапазон дат, когда виды запрещенных растений были замечены цветущими в Belwin Conservancy (A) и Carleton College (B).
Коренные виды прерий отмечены символом звездочки (*), а неродные виды отмечены символом плюса (+). Наблюдения проводились раз в две недели в течение всего периода, когда велась видеосъемка колоний медоносных пчел. Были определены и зарегистрированы все виды растений, цветущие в пятидесяти 1-метровых квадратах, расположенных случайным образом на пяти 200-метровых трансектах в восстановленных прериях на каждом участке. Поэтому, возможно, период цветения видов был более продолжительным, чем зафиксировано в ходе исследований.
(DOCX)
Ссылки
1.Attwood SJ, Maron M, House APN, Zammit C. Do arthropod assemblages display globally consistent responses to intensified agricultural land use and management? Glob Ecol Biogeogr. 2008;17: 585–599. https://doi.org/10.1111/j.1466-8238.2008.00399.x
View Article
Google Scholar
2.Hendrickx F, Maelfait J-P, Wingerden WV, Schweiger O, Speelmans M, Aviron S, et al. How landscape structure, land-use intensity and habitat diversity affect components of total arthropod diversity in agricultural landscapes. J Appl Ecol. 2007;44: 340–351. https://doi.org/10.1111/j.1365-2664.2006.01270.x
View Article
Google Scholar
3.Le Féon V, Schermann-Legionnet A, Delettre Y, Aviron S, Billeter R, Bugter R, et al. Intensification of agriculture, landscape composition and wild bee communities: A large scale study in four European countries. Agric Ecosyst Environ. 2010;137: 143–150. https://doi.org/10.1016/j.agee.2010.01.015
View Article
Google Scholar
4.Senapathi D, Goddard MA, Kunin WE, Baldock KCR. Landscape impacts on pollinator communities in temperate systems: evidence and knowledge gaps. Funct Ecol. 2017;31: 26–37. https://doi.org/10.1111/1365-2435.12809
View Article
Google Scholar
5.Kremen C, Williams NM, Thorp RW. Crop pollination from native bees at risk from agricultural intensification. Proc Natl Acad Sci. 2002;99: 16812–16816. https://doi.org/10.1073/pnas.262413599 pmid:12486221
View Article
PubMed/NCBI
Google Scholar
6.Potts SG, Biesmeijer JC, Kremen C, Neumann P, Schweiger O, Kunin WE. Global pollinator declines: Trends, impacts and drivers. Trends Ecol Evol. 2010;25: 345–353. https://doi.org/10.1016/j.tree.2010.01.007 pmid:20188434
View Article
PubMed/NCBI
Google Scholar
7.Morse R, Calderone NW. The value of honey bees as pollinators of U.S. Crops in 2000. Bee Cult. 2001;128: 1–15.
View Article
Google Scholar
8.Kulhanek K, Steinhauer N, Rennich K, Caron DM, Sagili RR, Pettis JS, et al. A national survey of managed honey bee 2015–2016 annual colony losses in the USA. J Apic Res. 2017;56: 328–340. https://doi.org/10.1080/00218839.2017.1344496
View Article
Google Scholar
9.Spivak M, Mader E, Vaughan M, Euliss NH. The plight of the bees. Environ Sci Technol. 2011;45: 34–38. https://doi.org/10.1021/es101468w pmid:20839858
View Article
PubMed/NCBI
Google Scholar
10.Goulson D, Nicholls E, Botías C, Rotheray EL. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science. 2015;347: 1255957. https://doi.org/10.1126/science.1255957 pmid:25721506
View Article
PubMed/NCBI
Google Scholar
11.Naug D. Nutritional stress due to habitat loss may explain recent honeybee colony collapses. Biol Conserv. 2009;142: 2369–2372. https://doi.org/10.1016/j.biocon.2009.04.007
View Article
Google Scholar
12.Otto CRV, Roth CL, Carlson BL, Smart MD. Land-use change reduces habitat suitability for supporting managed honey bee colonies in the Northern Great Plains. PNAS. 2016;113: 10430–10435. https://doi.org/10.1073/pnas.1603481113 pmid:27573824
View Article
PubMed/NCBI
Google Scholar
13.Smart M, Pettis J, Rice N, Browning Z, Spivak M. Linking measures of colony and individual honey bee health to survival among apiaries exposed to varying agricultural land use. PLOS ONE. 2016;11: e0152685. https://doi.org/10.1371/journal.pone.0152685 pmid:27027871
View Article
PubMed/NCBI
Google Scholar
14.Smart MD, Pettis JS, Euliss N, Spivak MS. Land use in the Northern Great Plains region of the U.S. influences the survival and productivity of honey bee colonies. Agric Ecosyst Environ. 2016;230: 139–149. https://doi.org/10.1016/j.agee.2016.05.030
View Article
Google Scholar
15.Brodschneider R, Crailsheim K. Nutrition and health in honey bees. Apidologie. 2010;41:278–294. https://doi.org/10.1051/apido/2010012
View Article
Google Scholar
16.Haydak MH. Honey bee nutrition. Annu Rev Entomol. 1970;15: 143–156. https://doi.org/10.1146/annurev.en.15.010170.001043
View Article
Google Scholar
17.Keller I, Fluri P, Imdorf A. Pollen nutrition and colony development in honey bees: part 1. Bee World. 2005;86: 3–10. https://doi.org/10.1080/0005772X.2005.11099641
View Article
Google Scholar
18.Scofield H, Mattila HR. Honey bee workers that are pollen stressed as larvae become poor foragers and waggle dancers as adults. PLOS ONE. 2015; 10(4): e0121731. https://doi.org/10.1371/journal.pone.0121731 pmid:25853902
View Article
PubMed/NCBI
Google Scholar
19.Huang Z. Honey bee nutrition. Am Bee J. 2010;150(8): 773–776
View Article
Google Scholar
20.Alaux C, Ducloz F, Crauser D, Le Conte Y. Diet effects on honeybee immunocompetence. Biol. Lett. 2010;6: 562–565. https://doi.org/10.1098/rsbl.2009.0986 pmid:20089536
View Article
PubMed/NCBI
Google Scholar
21.DeGrandi-Hoffman G, Chen Y, Rivera R, Carroll M, Chambers M, Hidalgo G, et al. Honey bee colonies provided with natural forage have lower pathogen loads and higher overwinter survival than those fed protein supplements. Apidologie. 2016;47: 186–196. https://doi.org/10.1007/s13592-015-0386-6
View Article
Google Scholar
22.Huang Z. Pollen nutrition affects honey bee stress resistance. Terr Arthropod Rev. 2012;5: 175–189. https://doi.org/10.1163/187498312X639568
View Article
Google Scholar
23.Pasquale GD, Salignon M, Conte YL, Belzunces LP, Decourtye A, Kretzschmar A, et al. Influence of pollen nutrition on honey bee health: do pollen quality and diversity matter? PLOS ONE. 2013;8: e72016. https://doi.org/10.1371/journal.pone.0072016 pmid:23940803
View Article
PubMed/NCBI
Google Scholar
24.Mao W, Schuler MA, Berenbaum MR. Honey constituents up-regulate detoxification and immunity genes in the western honey bee. PNAS. 2013;110(22): 8842–8846. https://doi.org/10.1073/pnas.1303884110 pmid:23630255
View Article
PubMed/NCBI
Google Scholar
25.Visscher PK, Seeley TD. Foraging strategy of honeybee colonies in a temperate deciduous forest. Ecology. 1982;63: 1790–1801.
View Article
Google Scholar
26.Seeley TD. The wisdom of the hive: The social physiology of honey bee colonies. Cambridge, MA: Harvard University Press; 1995.
27.Couvillon MJ, Fensome KA, Quah SK, Schürch R. Summertime blues. Commun Integr Biol. 2014;7: e28821. https://doi.org/10.4161/cib.28821 pmid:25346794
View Article
PubMed/NCBI
Google Scholar
28.Corby-Harris V, Bowsher J, Carr-markell MK, Carroll M, Centrella M, Cook SC, et al. Emerging themes from the ESA symposium entitled “Pollinator Nutrition: Lessons from Bees at Individual to Landscape Levels”. Bee World. 2019; 96(1): 3–9. https://doi.org/10.1080/0005772X.2018.1535951
View Article
Google Scholar
29.Garbuzov M, Ratnieks FLW. Listmania: The strengths and weaknesses of lists of garden plants to help pollinators. BioScience. 2014;64: 1019–1026. https://doi.org/10.1093/biosci/biu150
View Article
Google Scholar
30.MN BWSR. Identifying programs, funding and partner roles for Minnesota pollinator habitat. 2017. https://bwsr.state.mn.us/practices/pollinator/pollinator-toolbox.pdf
31.Wilson JS, Forister ML, Carril OM. Interest exceeds understanding in public support of bee conservation. Front Ecol Environ. 2017;15: 460–466. https://doi.org/10.1002/fee.1531
View Article
Google Scholar
32.Jordan WR, Lubick GM. Making nature whole: a history of ecological restoration. Washington D.C.: Island Press; 2011.
33.Weaver J. North American Prairie. Lincoln, NE: Johnsen Publishing; 1954.
34.Samson F, Knopf F. Prairie conservation in North America. BioScience. 1994;44: 418–421. https://doi.org/10.2307/1312365
View Article
Google Scholar
35.MN DNR. Minnesota Prairie Conservation Plan. 2011. https://www.dnr.state.mn.us/prairieplan/index.html
36.Couvillon MJ, Schürch R, Ratnieks FLW. Dancing bees communicate a foraging preference for rural lands in high-level agri-environment schemes. Curr Biol. 2014;24: 1212–1215. https://doi.org/10.1016/j.cub.2014.03.072 pmid:24856213
View Article
PubMed/NCBI
Google Scholar
37.Harmon-Threatt AN, Hendrix SD. Prairie restorations and bees: The potential ability of seed mixes to foster native bee communities. Basic Appl Ecol. 2015;16: 64–72. https://doi.org/10.1016/j.baae.2014.11.001
View Article
Google Scholar
38.Hopwood JL. The contribution of roadside grassland restorations to native bee conservation. Biol Conserv. 2008;141: 2632–2640. https://doi.org/10.1016/j.biocon.2008.07.026
View Article
Google Scholar
39.Tonietto RK, Ascher JS, Larkin DJ. Bee communities along a prairie restoration chronosequence: similar abundance and diversity, distinct composition. Ecol Appl. 2017;27: 705–717. https://doi.org/10.1002/eap.1481 pmid:27935661
View Article
PubMed/NCBI
Google Scholar
40.Beekman M, Ratnieks FLW. Long-range foraging by the honey-bee, Apis mellifera L. Funct Ecol. 2000;14: 490–496. https://doi.org/10.1046/j.1365-2435.2000.00443.x
View Article
Google Scholar
41.Couvillon MJ, Riddell Pearce FC, Accleton C, Fensome KA, Quah SKL, Taylor EL, et al. Honey bee foraging distance depends on month and forage type. Apidologie. 2015;46: 61–70. https://doi.org/10.1007/s13592-014-0302-5
View Article
Google Scholar
42.Härtel S, Steffan-Dewenter I. Ecology: Honey bee foraging in human-modified landscapes. Curr Biol. 2014;24: R524–R526. https://doi.org/10.1016/j.cub.2014.04.052 pmid:24892913
View Article
PubMed/NCBI
Google Scholar
43.Waddington KD, Herbert TJ, Visscher PK, Richter MR. Comparisons of forager distributions from matched honey bee colonies in suburban environments. Behav Ecol Sociobiol. 1994;35: 423–429. https://doi.org/10.1007/BF00165845
View Article
Google Scholar
44.von Frisch K. The Dance Language and Language and Orientation of Bees. Cambridge, MA: Harvard University Press; 1967.
45.Ratnieks FLW, Shackleton K. Does the waggle dance help honey bees to forage at greater distances than expected for their body size? Front Ecol Evol. 2015;3: 31.
View Article
Google Scholar
46.Seeley TD. Honey bee foragers as sensory units of their colonies. Behav Ecol Sociobiol. 1994;34: 51–62. https://doi.org/10.1007/BF00175458
View Article
Google Scholar
47.Horn J, Becher MA, Kennedy PJ, Osborne JL, Grimm V. Multiple stressors: using the honeybee model BEEHAVE to explore how spatial and temporal forage stress affects colony resilience. Oikos. 2016;125(7): 1001–1016.
View Article
Google Scholar
48.Bryant VM Jr, Jones GD. The r‐values of honey: Pollen coefficients, Palynology. 2001;25(1): 11–28. https://doi.org/10.1080/01916122.2001.9989554
View Article
Google Scholar
49.Jones GD. Pollen analyses for pollination research, acetolysis. J Pollinat Ecol. 2014;13: 203–217.
View Article
Google Scholar
50.Barron AB, Maleszka R, Meer RKV, Robinson GE. Octopamine modulates honey bee dance behavior. Proc Natl Acad Sci. 2007;104: 1703–1707. https://doi.org/10.1073/pnas.0610506104 pmid:17237217
View Article
PubMed/NCBI
Google Scholar
51.Jack-McCollough RT, Nieh JC. Honeybees tune excitatory and inhibitory recruitment signalling to resource value and predation risk. Anim Behav. 2015;110: 9–17. https://doi.org/10.1016/j.anbehav.2015.09.003
View Article
Google Scholar
52.Seeley TD, Mikheyev AS, Pagano GJ. Dancing bees tune both duration and rate of waggle-run production in relation to nectar-source profitability. J Comp Physiol A. 2000;186: 813–819. https://doi.org/10.1007/s003590000134 pmid:11085635
View Article
PubMed/NCBI
Google Scholar
53.Beekman M, Preece K, Schaerf TM. Dancing for their supper: Do honeybees adjust their recruitment dance in response to the protein content of pollen? Insectes Sociaux. 2016;63: 117–126. https://doi.org/10.1007/s00040-015-0443-1
View Article
Google Scholar
54.Zarchin S, Dag A, Salomon M, Hendriksma HP, Shafir S. Honey bees dance faster for pollen that complements colony essential fatty acid deficiency. Behav Ecol Sociobiol. 2017;71: 172. https://doi.org/10.1007/s00265-017-2394-1
View Article
Google Scholar
55.Couvillon MJ, Schürch R, Ratnieks FLW. Waggle dance distances as integrative indicators of seasonal foraging challenges. PLOS ONE. 2014;9: e93495. https://doi.org/10.1371/journal.pone.0093495 pmid:24695678
View Article
PubMed/NCBI
Google Scholar
56.Schürch R, Couvillon MJ, Burns DDR, Tasman K, Waxman D, Ratnieks FLW. Incorporating variability in honey bee waggle dance decoding improves the mapping of communicated resource locations. J Comp Physiol A. 2013;199: 1143–1152. https://doi.org/10.1007/s00359-013-0860-4
View Article
Google Scholar
57.ESRI 2019. ArcGIS Desktop: Release 10.7.1. Redlands, CA: Environmental Systems Research Institute.
58.Langstroth LL. Langstroth on the Hive and Honey Bee: A Beekeeper’s Manual. Northampton: Hopkins, Bridgman, and Company; 1853.
59.Corrigan CE. Relationship between relative hive entrance position and dance floor location [undergraduate honors thesis]. Johnson City (TN): East Tennessee State University; 2014.
60.Robinson GE, Underwood BA, Henderson CE. A highly specialized water-collecting honey bee. Apidologie. 1984;15: 355–358. https://doi.org/10.1051/apido:19840307
View Article
Google Scholar
61.Meyer W, Ulrich W. “Propolis bees” and their activities. Bee World. 1956;37: 25–36. https://doi.org/10.1080/0005772X.1956.11094916
View Article
Google Scholar
62.Nakamura J, Seeley TD. The functional organization of resin work in honeybee colonies. Behav Ecol Sociobiol. 2006;60: 339–349. https://doi.org/10.1007/s00265-006-0170-8
View Article Google Scholar
63.Simone-Finstrom MD, Spivak M. Increased resin collection after parasite challenge: a case of self-medication in honey bees? PLOS ONE. 2012;7: e34601. https://doi.org/10.1371/journal.pone.0034601 pmid:22479650
View Article
PubMed/NCBI
Google Scholar
64.Couvillon MJ, Pearce FCR, Harris-Jones EL, Kuepfer AM, Mackenzie-Smith SJ, Rozario LA, Schürch R, Ratnieks FLW. Intra-dance variation among waggle runs and the design of efficient protocols for honey bee dance decoding. Biol Open. 2012;BIO20121099. https://doi.org/10.1242/bio.20121099
View Article
Google Scholar
65.Hodges D. The pollen loads of the honey bee: a guide to their identification by colour and form. London: Bee Research Institute Limited; 1984.
66.Free JB. The flower constancy of honeybees. J Anim Ecol. 1963;32: 119–131. https://doi.org/10.2307/2521
View Article
Google Scholar
67.Hernández DL, Esch EH, Alster CJ, McKone MJ, Camill P. Rapid accumulation of soil carbon and nitrogen in a prairie restoration chronosequence. Soil Sci Soc Am J. 2013;77: 2029–2038. https://doi.org/10.2136/sssaj2012.0403
View Article
Google Scholar
68.Cornman RS, Otto CRV, Iwanowicz D, Pettis JS. Taxonomic Characterization of honey bee (Apis mellifera) pollen foraging based on non-overlapping paired-end sequencing of nuclear ribosomal loci. PLOS ONE. 2015;10: e0145365. https://doi.org/10.1371/journal.pone.0145365 pmid:26700168
View Article
PubMed/NCBI
Google Scholar
69.Richardson RT, Lin CH, Quijia JO, Riusech NS, Goodell K, Johnson RM. Rank-based characterization of pollen assemblages collected by honey bees using a multi-locus metabarcoding approach. Appl Plant Sci. 2015;3: 1500043. https://doi.org/10.3732/apps.1500043
View Article
Google Scholar
70.Brown LD, Cai TT, DasGupta A. Interval estimation for a binomial proportion. Stat Sci. 2001;16: 101–117.
View Article
Google Scholar
71.Garbuzov M, Couvillon MJ, Schürch R, Ratnieks FLW. Honey bee dance decoding and pollen-load analysis show limited foraging on spring-flowering oilseed rape, a potential source of neonicotinoid contamination. Agric Ecosyst Environ. 2015;203: 62–68. https://doi.org/10.1016/j.agee.2014.12.009
View Article
Google Scholar
72.Seeley TD. Social foraging in honey bees: how nectar foragers assess their colony’s nutritional status. Behav Ecol Sociobiol. 1989;24(3): 181–199. https://doi.org/10.1007/BF00292101
View Article
Google Scholar
73.Tanner D, Visscher K. Does the body orientation of waggle dance followers affect the accuracy of recruitment? Apidologie. 2009;40: 55–62. https://doi.org/10.1051/apido:2008074
View Article
Google Scholar
74.Seeley TD, Camazine S, Sneyd J. Collective decision-making in honey bees: how colonies choose among nectar sources. Behav Ecol Sociobiol. 1991;28: 277. https://doi.org/10.1007/BF00175101
75.Sponsler DB, Matcham EG, Lin C-H, Lanterman JL, Johnson RM. Spatial and taxonomic patterns of honey bee foraging: A choice test between urban and agricultural landscapes. J Urban Ecol. 2017;3(1): juw008. https://doi.org/10.1093/jue/juw008
View Article
Google Scholar
76.Smart MD, Cornman RS, Iwanowicz DD, McDermott-Kubeczko M, Pettis JS, Spivak MS, et al. A comparison of honey bee-collected pollen from working agricultural lands using light microscopy and its metabarcoding. Environ Entomol. 2017;46: 38–49. https://doi.org/10.1093/ee/nvw159 pmid:28062536
View Article
PubMed/NCBI
Google Scholar
77.Wood TJ, Kaplan I, Szendrei Z. Wild bee pollen diets reveal patterns of seasonal foraging resources for honey bees. Front Ecol Evol. 2018;6: 210. https://doi.org/10.3389/fevo.2018.00210
View Article
Google Scholar
78.Dolezal AG, St. Clair AL, Zhang G, Toth AL, O’Neal ME. Native habitat mitigates feast–famine conditions faced by honey bees in an agricultural landscape. PNAS. 2019; 201912801. https://doi.org/10.1073/pnas.1912801116
79.Furgala B, Spivak M, Reuter GS. Beekeeping in Northern Climates. St. Paul: University of Minnesota; 2017.
80.Haydak MH. Beekeeping in Minnesota. St. Paul: University of Minnesota; 1962.
81.Hovenden MJ, Wills KE, Schoor JKV, Williams AL, Newton PCD. Flowering phenology in a species-rich temperate grassland is sensitive to warming but not elevated CO2. New Phytol. 2008;178: 815–822. https://doi.org/10.1111/j.1469-8137.2008.02419.x pmid:18346104
View Article
PubMed/NCBI
Google Scholar
82.Lesica P, Kittelson PM. Precipitation and temperature are associated with advanced flowering phenology in a semi-arid grassland. J Arid Environ. 2010;74: 1013–1017. https://doi.org/10.1016/j.jaridenv.2010.02.002
View Article
Google Scholar
83.Scaven VL, Rafferty NE. Physiological effects of climate warming on flowering plants and insect pollinators and potential consequences for their interactions. Curr Zool. 2013;59: 418–426. https://doi.org/10.1093/czoolo/59.3.418 pmid:24009624
View Article
PubMed/NCBI
Google Scholar
84.Werner PA, Gross RS, Bradbury IK. The biology of Canadian weeds: 45 Solidago canadensis L. Can J Plant Sci. 1980;60: 1393–1409. https://doi.org/10.4141/cjps80-194
View Article
Google Scholar
85.Beuselinck PR, Steiner JJ. Birdsfoot trefoil origins and germplasm diversity. In: Beuselinck PR, editor. Trefoil: The Science and Technology of Lotus. CSSA Special Publication no 28. Madison, WI: CSSA and ASA; 1999, p. 81–96. https://doi.org/10.2135/cssaspecpub28.c5
86.Hung K-LJ, Kingston JM, Albrecht M, Holway DA, Kohn JR. The worldwide importance of honey bees as pollinators in natural habitats. Proc Roy Acad Sci B. 2018;285: 1870. https://doi.org/10.1098/rspb.2017.2140
View Article
Google Scholar
87.Mallinger RE, Gaines-Day HR, Gratton C. Do managed bees have negative effects on wild bees?: A systematic review of the literature. PLoS ONE. 2017;12(12): e0189268. https://doi.org/10.1371/journal.pone.0189268 pmid:29220412
View Article
PubMed/NCBI
Google Scholar
88.Swenk MH. Two new bees of the genus Colletes, oligotropic upon Petalostemum. Am. Mus. Novit. 1925;186: 1–9.
View Article
Google Scholar
Добавить комментарий